ADN
Definición

El ácido desoxirribonucleico (/ diɒksɪraʊboʊnjuːkliːɪk, -kleɪ- / (

Las dos cadenas de ADN también se conocen como polinucleótidos ya que están compuestas de unidades monoméricas más simples llamadas nucleótidos. Cada nucleótido se compone de una de cuatro nucleobases que contienen nitrógeno (citosina [C], guanina [G], adenina [A] o timina [T]), un azúcar llamado desoxirribosa y un grupo fosfato. Los nucleótidos están unidos entre sí en una cadena por enlaces covalentes entre el azúcar de un nucleótido y el fosfato del siguiente, lo que da como resultado una cadena principal de azúcar-fosfato alternante. Las bases nitrogenadas de las dos cadenas de polinucleótidos separadas están unidas entre sí, de acuerdo con las reglas de emparejamiento de bases (A con T y C con G), con enlaces de hidrógeno para formar ADN bicatenario.
Las bases nitrogenadas complementarias se dividen en dos grupos, pirimidinas y purinas. En el ADN, las pirimidinas son timina y citosina; las purinas son adenina y guanina.
El ADN almacena información biológica. La cadena principal del ADN es resistente a la escisión, y ambas cadenas de la estructura bicatenaria almacenan la misma información biológica. Esta información se replica a medida que los dos filamentos se separan. Una gran parte del ADN (más del 98% para humanos) no codifica, lo que significa que estas secciones no sirven como patrones para las secuencias de proteínas.
Las dos cadenas de ADN corren en direcciones opuestas entre sí y, por lo tanto, son antiparalelas. Se adjunta a cada azúcar uno de los cuatro tipos de nucleobases (informalmente, bases ). Es la secuencia de estas cuatro nucleobases a lo largo de la cadena principal que codifica la información genética. Las cadenas de ARN se crean usando cadenas de ADN como plantilla en un proceso llamado transcripción. Bajo el código genético, estos RNAstrands se traducen para especificar la secuencia de aminoácidos dentro de las proteínas en un proceso llamado traducción.
Dentro de las células eucariotas, el ADN se organiza en estructuras largas llamadas cromosomas. Antes de la división celular típica, estos cromosomas se duplican en el proceso de replicación del ADN, proporcionando un conjunto completo de cromosomas para cada célula hija. Los organismos eucarióticos (animales, plantas, hongos y protistas) almacenan la mayor parte de su ADN dentro del núcleo celular y parte de su ADN en orgánulos, como las mitocondrias o los cloroplastos. Por el contrario, los procariotas (bacterias y arqueas) almacenan su ADN solo en el citoplasma. Dentro de los cromosomas eucariotas, las proteínas de la cromatina, como las histonas, compactas y organizan el ADN. Estas estructuras compactas guían las interacciones entre el ADN y otras proteínas, ayudando a controlar qué partes del ADN se transcriben.
El ADN fue aislado primero por Friedrich Miescher en 1869. Su estructura molecular fue identificada por primera vez por James Watson y Francis Crick en el Laboratorio Cavendish de la Universidad de Cambridge en 1953, cuyos esfuerzos de construcción de modelos fueron guiados por datos de difracción de rayos X adquiridos por Raymond Gosling, quien era un estudiante de posgrado de Rosalind Franklin. El ADN es utilizado por los investigadores como una herramienta molecular para explorar leyes y teorías físicas, como el teorema ergódico y la teoría de la elasticidad. Las propiedades materiales únicas del ADN lo han convertido en una molécula atractiva para los científicos e ingenieros de materiales interesados en la micro y nanofabricación. Entre los avances notables en este campo están el origami de ADN y los materiales híbridos basados en ADN.
Propiedades

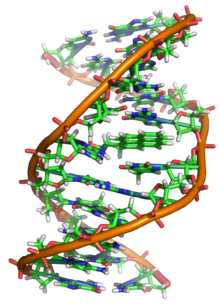
El ADN es un polímero largo hecho de unidades repetitivas llamadas nucleótidos. La estructura del ADN es dinámica a lo largo de su longitud, siendo capaz de enrollarse en bucles apretados y otras formas. En todas las especies se compone de dos cadenas helicoidales, unidas entre sí por enlaces de hidrógeno. Ambas cadenas están enrolladas alrededor del mismo eje, y tienen el mismo paso de 34 ångströms (3.4 nanómetros). El par de cadenas tiene un radio de 10 ångströms (1.0 nanómetro). Según otro estudio, cuando se midió en una solución diferente, la cadena de ADN midió de 22 a 26 ångströms de ancho (2,2 a 2,6 nanómetros), y una unidad de nucleótidos midió 3,3 Å (0,33 nm) de largo. Aunque cada unidad de repetición de nucleótidos individual es muy pequeña, los polímeros de ADN pueden ser moléculas muy grandes que contienen de millones a cientos de millones de nucleótidos. Por ejemplo, el ADN en el cromosoma humano más grande,
En los organismos vivos, el ADN no suele existir como una sola molécula, sino como un par de moléculas que se mantienen unidas. Estas dos largas hebras se entrelazan como vides, en forma de doble hélice. El nucleótido contiene un segmento de la columna vertebral de la molécula (que mantiene la cadena unida) y una nucleobase (que interactúa con la otra cadena de ADN en la hélice). Una nucleobase vinculada a un azúcar se llama nucleósido y una base ligada a un azúcar y uno o más grupos de fosfato se llama nucleótido. Un polímero que comprende múltiples nucleótidos unidos (como en el ADN) se llama un polinucleótido.
La columna vertebral de la cadena de ADN está hecha de fosfato y residuos de azúcar alternados. El azúcar en el ADN es 2-desoxirribosa, que es un azúcar pentosa (cinco carbonos). Los azúcares se unen por grupos de fosfato que forman enlaces fosfodiéster entre el tercero y el quinto átomo de carbono de los anillos de azúcar adyacentes, que se conocen como los carbonos 3 'y 5', el símbolo principal se utiliza para distinguir estos átomos de carbono de los del base a la cual la desoxirribosa forma un enlace glucosídico. Al imaginar el ADN, normalmente se considera que cada fosforilo "pertenece" al nucleótido cuyo carbono 5 'forma un enlace con el mismo. Por lo tanto, cualquier cadena de ADN normalmente tiene un extremo en el que hay un fosforilo unido al carbono 5 'de una ribosa (el 5' fosforilo) y otro extremo en el que hay un hidroxilo libre unido al carbono 3 'de una ribosa (el 3 'hidroxilo). La orientación de los carbonos 3 'y 5' a lo largo de la cadena principal de azúcar-fosfato confiere direccionalidad (a veces llamada polaridad) a cada cadena de ADN. En una doble hélice, la dirección de los nucleótidos en una cadena es opuesta a su dirección en la otra cadena: las cadenas son antiparalelo . Se dice que los extremos asimétricos de las cadenas de ADN tienen una direccionalidad de cinco primos (5 ') y tres primos (3'), con el extremo 5 'que tiene un grupo fosfato terminal y el extremo 3' un grupo hidroxilo terminal. Una diferencia importante entre el ADN y el ARN es el azúcar, con la 2-desoxirribosa en el ADN reemplazada por la pentosa azúcar ribosa alternativa en el ARN.
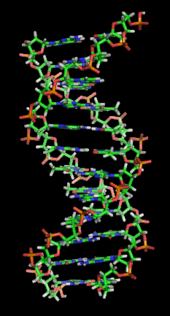
La doble hélice de ADN se estabiliza principalmente por dos fuerzas: enlaces de hidrógeno entre nucleótidos e interacciones de apilamiento de bases entre nucleobases aromáticos. En el entorno acuoso de la célula, los enlaces
π conjugados de las bases de nucleótidos se alinean perpendicularmente al eje de la molécula de ADN, minimizando su interacción con la capa de solvatación. Las cuatro bases encontradas en el ADN son adenina (A), citosina (C), guanina (G) y timina (T). Estas cuatro bases están unidas al azúcar-fosfato para formar el nucleótido completo, como se muestra para el monofosfato de adenosina. Adenine se empareja con timina y pares de guanina con citosina. Fue representado por pares de bases AT y pares de bases GC.
π conjugados de las bases de nucleótidos se alinean perpendicularmente al eje de la molécula de ADN, minimizando su interacción con la capa de solvatación. Las cuatro bases encontradas en el ADN son adenina (A), citosina (C), guanina (G) y timina (T). Estas cuatro bases están unidas al azúcar-fosfato para formar el nucleótido completo, como se muestra para el monofosfato de adenosina. Adenine se empareja con timina y pares de guanina con citosina. Fue representado por pares de bases AT y pares de bases GC.
Clasificación de nucleobases
Las nucleobases se clasifican en dos tipos: las purinas, A y G, que son compuestos heterocíclicos de cinco y seis miembros fusionados, y las pirimidinas, los anillos C y T. de seis miembros. Una quinta base de pirimidina, el uracilo (U), por lo general toma el lugar de la timina en el ARN y se diferencia de la timina por la falta de un grupo metilo en su anillo. Además del ARN y el ADN, se han creado muchos análogos de ácidos nucleicos artificiales para estudiar las propiedades de los ácidos nucleicos o para su uso en biotecnología.
Bases no canónicas
El uracilo no se encuentra generalmente en el ADN, y se presenta solo como un producto de degradación de la citosina. Sin embargo, en varios bacteriófagos, Bacillus subtilis bacteriófagos PBS1 y PBS2 y Yersinia bacteriófago piR1-37, la timina ha sido reemplazada por uracilo. Otro fago, el Staphylococcal fago S6, ha sido identificado con un genoma en el que la timina ha sido reemplazada por uracilo.
La 5-hidroximetildesoxiuridina, (hm5dU) también es conocida por reemplazar la timidina en varios genomas, incluidos los Bacillus phages SPO1, φe, SP8, H1, 2C y SP82. Otro uracilo modificado, 5-dihidroxipentauracilo, también se ha descrito.
Base J (beta-d-glucopiranosiloximetiluracilo), una forma modificada de uracilo, también se encuentra en varios organismos: los flagelados Diplonema y Euglena , y todos los géneros de cinetoplastia. La síntesis de J se produce en dos pasos: en el primer paso, un específico la timidina en el ADN se convierte en hidroximetilodesoxiuridina; en el segundo, HOMedU está glicosilado para formar J. Se han identificado proteinas que se unen específicamente a esta base. Estas proteínas parecen ser parientes distantes del oncogén Tet1 que está involucrado en la patogénesis de la leucemia mieloide aguda. J parece actuar como una señal de terminación para la ARN polimerasa II.
En 1976, se encontró que un bacteriófago - S-2L - que infecta especies del género Synechocystistiene todas las bases de adenosina dentro de su genoma reemplazadas por 2,6-diaminopurina. En 2016 se descubrió que la desoxirchaeosina estaba presente en los genomas de varias bacterias y el fago Escherichia 9g.
Las bases modificadas también ocurren en el ADN. El primero de estos reconocidos fue 5-metilcitosina que se encontró en el genoma de Mycobacterium tuberculosis en 1925. El reemplazo completo de citosina por 5-glicosilhidroximetilcitosina en T incluso los fagos (T2, T4 y T6) se observó en 1953 en los genomas de Xanthomonas oryzae bacteriófago Xp12 y halovirus FH, el complemento completo de cistosina ha sido reemplazado por 5-metilcitosina. Se descubrió que la 6N-metiadenina estaba presente en el ADN en 1955. La N6-carbamoil-metiladenina se describió en 1975. La 7-metilguanina se describió en 1976. La N4-metilcitosina en el ADN se describió en 1983. En 1985 se encontró 5-hidroxicitosina en el genomas de Rhizobium phages RL38JI y N17. α-putrescinylthymine se produce en los dos genomas de Delftia fago ΦW-14 y Bacillus phage SP10. La α-glutamiltimidina se encuentra en el fago Bacillus SP01 y el 5-dihidroxypentiluracilo se encuentra en el fago Bacillus SP15.
Se desconoce el motivo de la presencia de estas bases no canónicas en el ADN. Parece probable que al menos parte de la razón de su presencia en virus bacterianos (fagos) es evitar las enzimas de restricción presentes en las bacterias. Este sistema enzimático actúa al menos en parte como un sistema inmunitario molecular que protege a las bacterias de la infección por virus.
Esta no parece ser la historia completa. Se han informado cuatro modificaciones a los residuos de citosina en el ADN humano. Estas modificaciones son la adición de metilo (CH 3 ) -, hidroximetilo (CH 2 OH) -, formilo (CHO) - y carboxilo (COOH) - grupos. Se cree que estas modificaciones tienen funciones regulatorias.
El uracilo se encuentra en las regiones centroméricas de al menos dos cromosomas humanos (6 y 11).
Listado de bases no canónicas encontradas en el ADN
Se sabe que hay diecisiete bases no canónicas en el ADN. La mayoría de estos son modificaciones de las bases canónicas más uracilo.
- Adenosina modificada
- N6-carbamoil-metiladenina
- N6-methyadenine
- Guanine modificado
- 7-metilguanina
- Citosina modificada
- N4-Methylcytosine
- 5-Carboxylcytosine
- 5-formilcitosina
- 5-glicosilhidroximetilcitosina
- 5-Hidroxicitosina
- 5-metilcitosina
- Timidina modificada
- α-Glutamythymidine
- α-Putrescinylthymine
- Uracil y modificaciones
- Base J
- Uracil
- 5-Dihydroxypentauracil
- 5-Hydroxymethyldeoxyuracil
- Otros
- Deoxyarchaeosine
- 2,6-Diaminopurina

Surcos
Las hebras helicoidales gemelas forman la columna vertebral del ADN. Se puede encontrar otra doble hélice trazando los espacios, o surcos, entre los filamentos. Estos huecos son adyacentes a los pares de bases y pueden proporcionar un sitio de unión. Como los hilos no están situados simétricamente uno con respecto al otro, las ranuras tienen un tamaño desigual. Una ranura, la ranura principal, tiene 22 Å de ancho y la otra, la ranura menor, tiene 12 Å de ancho. El ancho de la ranura principal significa que los bordes de las bases son más accesibles en la ranura principal que en la ranura menor. Como resultado, las proteínas como los factores de transcripción que pueden unirse a secuencias específicas en el ADN bicatenario normalmente hacen contacto con los lados de las bases expuestas en el surco mayor. Esta situación varía en conformaciones inusuales de ADN dentro de la célula (ver más abajo), pero los surcos mayor y menor siempre se nombran para reflejar las diferencias de tamaño que se verían si el ADN se retuerce en la forma B ordinaria.
Emparejamiento de bases
En una doble hélice de ADN, cada tipo de nucleobase en una cadena se une con un solo tipo de nucleobase en la otra cadena. Esto se llama emparejamiento de bases complementarias. Aquí, las purinas forman enlaces de hidrógeno con las pirimidinas, con la adenina que se une solo a la timina en dos enlaces de hidrógeno, y la citosina que se une solo a la guanina en tres enlaces de hidrógeno. Esta disposición de dos nucleótidos que se unen a través de la doble hélice se llama un par de bases Watson-Crick. Otro tipo de emparejamiento de bases es el apareamiento de bases de Hoogsteen donde se forman dos enlaces de hidrógeno entre la guanina y la citosina. Como los enlaces de hidrógeno no son covalentes, se pueden romper y volver a unir con relativa facilidad. Las dos hebras de ADN en una doble hélice se pueden separar como una cremallera, ya sea por una fuerza mecánica o alta temperatura. Como resultado de esta complementariedad de pares de bases, toda la información en la secuencia bicatenaria de una hélice de ADN se duplica en cada cadena, que es vital en la replicación del ADN. Esta interacción reversible y específica entre pares de bases complementarios es crítica para todas las funciones del ADN en organismos vivos.
 |
 |
Top, un par de bases GC con tres enlaces de hidrógeno. Abajo, un par base AT con dos enlaces de hidrógeno. Los enlaces de hidrógeno no covalentes entre los pares se muestran como líneas discontinuas.
Los dos tipos de pares de bases forman diferentes números de enlaces de hidrógeno, AT forma dos enlaces de hidrógeno y GC forma tres enlaces de hidrógeno (ver figuras, derecha). El ADN con alto contenido de GC es más estable que el ADN con bajo contenido de GC.
Como se señaló anteriormente, la mayoría de las moléculas de ADN son en realidad dos cadenas de polímeros, unidas de forma helicoidal por enlaces no covalentes; esta estructura bicatenaria ( dsDNA ) se mantiene en gran medida mediante interacciones de apilamiento de bases intrastrand, que son más fuertes para las acumulaciones G, C. Los dos filamentos pueden separarse, un proceso conocido como fusión, para formar dos moléculas de ADN monocatenario ( ssDNA ). La fusión se produce a alta temperatura, bajo contenido de sal y pH alto (el pH bajo también derrite el ADN, pero dado que el ADN es inestable debido a la depuración con ácido, rara vez se usa un pH bajo).
La estabilidad de la forma de dsDNA depende no solo del contenido de GC (pares de base% G, C) sino también de la secuencia (dado que el apilamiento es específico de la secuencia) y también de la longitud (las moléculas más largas son más estables). La estabilidad se puede medir de varias maneras; una forma común es la "temperatura de fusión", que es la temperatura a la que el 50% de las moléculas de ds se convierten en moléculas ss; la temperatura de fusión depende de la fuerza iónica y la concentración de ADN. Como resultado, es tanto el porcentaje de pares de bases de GC como la longitud total de una doble hélice de ADN lo que determina la fuerza de la asociación entre las dos cadenas de ADN. Las largas hélices de ADN con un alto contenido de GC tienen cadenas que interactúan más fuertes, mientras que las hélices cortas con un alto contenido de AT tienen cadenas que interactúan más débilmente. En biología,
En el laboratorio, la fuerza de esta interacción se puede medir al encontrar la temperatura necesaria para romper los enlaces de hidrógeno, su temperatura de fusión (también llamada valor de T m ). Cuando todos los pares de bases en una doble hélice de ADN se derriten, los filamentos se separan y existen en solución como dos moléculas totalmente independientes. Estas moléculas de ADN monocatenario no tienen una única forma común, pero algunas conformaciones son más estables que otras.
Sentido y antisentido
Una secuencia de ADN se denomina "sentido" si su secuencia es la misma que la de una copia de ARN mensajero que se traduce en proteína. La secuencia en la cadena opuesta se denomina secuencia "antisentido". Ambas secuencias sentido y antisentido pueden existir en diferentes partes de la misma cadena de ADN (es decir, ambas cadenas pueden contener secuencias sentido y antisentido). Tanto en procariotas como en eucariotas, se producen secuencias de ARN antisentido, pero las funciones de estos ARN no son del todo claras. Una propuesta es que los ARNs antisentido están implicados en la regulación de la expresión génica a través del emparejamiento de bases ARN-ARN.
Algunas secuencias de ADN en procariotas y eucariotas, y más en plásmidos y virus, difuminan la distinción entre cadenas sentido y antisentido al tener genes superpuestos. En estos casos, algunas secuencias de ADN tienen una doble función, codificar una proteína cuando se lee a lo largo de una cadena, y una segunda proteína cuando se lee en la dirección opuesta a lo largo de la otra cadena. En las bacterias, esta superposición puede estar involucrada en la regulación de la transcripción génica, mientras que en los virus, la superposición de genes aumenta la cantidad de información que puede codificarse dentro del genoma viral pequeño.
Supercoiling
El ADN se puede retorcer como una cuerda en un proceso llamado superenrollamiento de ADN. Con el ADN en su estado "relajado", una hebra generalmente circula alrededor del eje de la doble hélice una vez cada 10,4 pares de bases, pero si el ADN se retuerce, los filamentos se vuelven más apretados o más sueltos. Si el ADN se retuerce en la dirección de la hélice, este es un superenrrollamiento positivo, y las bases se mantienen más unidas. Si se tuercen en la dirección opuesta, esto es supercoil negativo, y las bases se separan más fácilmente. En la naturaleza, la mayoría del ADN tiene un ligero superenrrollamiento negativo que es introducido por las enzimas llamadas topoisomerasas. Estas enzimas también son necesarias para aliviar las tensiones de torsión introducidas en las cadenas de ADN durante procesos tales como la transcripción y la replicación del ADN.

Estructuras de ADN alternativas
El ADN existe en muchas conformaciones posibles que incluyen las formas A-DNA, B-DNA y Z-DNA, aunque solo se han observado directamente B-DNA y Z-DNA en organismos funcionales. La conformación que adopta el ADN depende del nivel de hidratación, la secuencia de ADN, la cantidad y dirección de superenrrollamiento, las modificaciones químicas de las bases, el tipo y la concentración de iones metálicos y la presencia de poliaminas en solución.
Los primeros informes publicados de patrones de difracción de rayos X de A-DNA, y también análisis de B-DNA utilizados basados en transformaciones de Patterson que proporcionaron solo una cantidad limitada de información estructural para fibras orientadas de DNA. Un análisis alternativo fue propuesto por Wilkins et al. , en 1953, para los patrones de dispersión de difracción de rayos X de ADN-B in vivo de fibras de ADN altamente hidratadas en términos de cuadrados de funciones de Bessel. En la misma revista, James Watson y Francis Crick presentaron su análisis de modelado molecular de los patrones de difracción de rayos X del ADN para sugerir que la estructura era una doble hélice.
Aunque la forma de B-DNA es más común en las condiciones que se encuentran en las células, no es una conformación bien definida, sino una familia de conformaciones de ADN relacionadas que se producen a los altos niveles de hidratación presentes en las células vivas. Sus correspondientes patrones de difracción y dispersión de rayos X son característicos de los paracristales moleculares con un grado significativo de desorden.
En comparación con el B-DNA, la forma de A-DNA es una espiral derecha más ancha, con un groove menor amplio y poco profundo y un groove mayor más estrecho y profundo. La forma A se produce en condiciones no fisiológicas en muestras de ADN parcialmente deshidratadas, mientras que en la célula se puede producir en pares híbridos de cadenas de ADN y ARN, y en complejos enzima-ADN. Los segmentos de ADN en los que las bases se han modificado químicamente por metilación pueden experimentar un cambio mayor en la conformación y adoptar la forma Z. Aquí, los hilos giran alrededor del eje helicoidal en una espiral hacia la izquierda, lo opuesto a la forma B más común. Estas estructuras inusuales pueden ser reconocidas por proteínas de unión de Z-DNA específicas y pueden estar involucradas en la regulación de la transcripción.
Química de ADN alternativa
Durante muchos años, los exobiólogos han propuesto la existencia de una biosfera sombría, una biosfera microbiana postulada de la Tierra que utiliza procesos bioquímicos y moleculares radicalmente diferentes a los de la vida actualmente conocida. Una de las propuestas fue la existencia de formas de vida que usan arsénico en lugar de fósforo en el ADN. Un informe en 2010 de la posibilidad en la bacteria GFAJ-1, fue anunciado, aunque la investigación fue disputada, y la evidencia sugiere que la bacteria activamente evita la incorporación de arsénico en la estructura del ADN y otras biomoléculas.
Estructuras Quadruplex
En los extremos de los cromosomas lineales hay regiones especializadas de ADN llamadas telómeros. La función principal de estas regiones es permitir que la célula replique los extremos del cromosoma utilizando la enzima telomerasa, ya que las enzimas que normalmente replican el ADN no pueden copiar los extremos 3 'de los cromosomas. Estas tapas cromosómicas especializadas también ayudan a proteger los extremos del ADN y evitan que los sistemas de reparación del ADN en la célula los traten como un daño que debe corregirse. En células humanas, los telómeros son generalmente longitudes de ADN monocatenario que contiene varios miles de repeticiones de una secuencia TTAGGG simple.
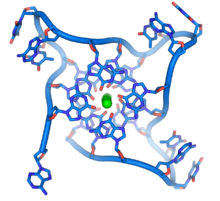
Estas secuencias ricas en guanina pueden estabilizar los extremos de los cromosomas formando estructuras de conjuntos apilados de unidades de cuatro bases, en lugar de los pares de bases habituales encontrados en otras moléculas de ADN. Aquí, cuatro bases de guanina forman una placa plana y estas unidades planas de cuatro bases se apilan una encima de la otra para formar una estructura G-quadruplex estable. Estas estructuras se estabilizan mediante enlaces de hidrógeno entre los bordes de las bases y la quelación de un ion metálico en el centro de cada unidad de cuatro bases. También se pueden formar otras estructuras, con el conjunto central de cuatro bases que provienen de una sola hebra doblada alrededor de las bases o de varias hebras paralelas diferentes, cada una de las cuales contribuye con una base a la estructura central.
Además de estas estructuras apiladas, los telómeros también forman estructuras de bucles grandes llamadas bucles de telómero o bucles en T. Aquí, el ADN monocatenario se riza en un círculo largo estabilizado por proteínas que se unen a los telómeros. Al final del ciclo en T, el ADN del telómero monocatenario se mantiene en una región del ADN bicatenario por la cadena del telómero, lo que altera el ADN de doble hélice y el emparejamiento de bases con una de las dos cadenas. Esta estructura de triple cadena se denomina bucle de desplazamiento o D-loop.
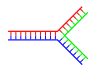 |  |
| Una rama | Varias ramas |
El ADN ramificado puede formar redes que contienen ramas múltiples.
ADN ramificado
En el ADN, se produce deshilachado cuando existen regiones no complementarias al final de una cadena doble de ADN complementaria. Sin embargo, el ADN ramificado puede ocurrir si se introduce una tercera cadena de ADN y contiene regiones adyacentes capaces de hibridarse con las regiones deshilachadas de la cadena doble preexistente. Aunque el ejemplo más simple de ADN ramificado implica solo tres cadenas de ADN, también son posibles complejos que involucran cadenas adicionales y ramas múltiples. El ADN ramificado se puede utilizar en nanotecnología para construir formas geométricas, ver la sección sobre usos en la tecnología a continuación.
Modificaciones químicas y embalaje de ADN alterado
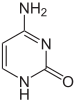 |  |  |
| citosina | 5-metilcitosina | timina |
Estructura de la citosina con y sin el grupo 5-metilo. La desaminación convierte 5-metilcitosina en timina.
Modificaciones de base y empaque de ADN
La expresión de los genes está influenciada por la forma en que el ADN se empaqueta en los cromosomas, en una estructura llamada cromatina. Las modificaciones de la base pueden estar involucradas en el empaquetamiento, con regiones que tienen baja o nula expresión génica que generalmente contienen altos niveles de metilación de bases de citosina. El empaquetamiento del ADN y su influencia en la expresión génica también pueden ocurrir por modificaciones covalentes del núcleo de proteína de histona alrededor del cual el ADN se envuelve en la estructura de cromatina o por remodelación llevada a cabo por complejos de remodelación de cromatina (ver remodelación de cromatina). Existe, además, diafonía entre la metilación del ADN y la modificación de histonas, por lo que pueden afectar coordinadamente a la cromatina y la expresión génica.
Por ejemplo, la metilación de citosina produce 5-metilcitosina, que es importante para la inactivación de X de los cromosomas. El nivel promedio de metilación varía entre los organismos: el gusano Caenorhabditis elegans carece de metilación de la citosina, mientras que los vertebrados tienen niveles más altos, con hasta un 1% de su ADN que contiene 5-metilcitosina. A pesar de la importancia de la 5-metilcitosina, puede desaminarse para dejar una base de timina, por lo que las citosinas metiladas son particularmente propensas a las mutaciones. Otras modificaciones de base incluyen la metilación de adenina en bacterias, la presencia de 5-hidroximetilcitosina en el cerebro y la glicosilación de uracilo para producir la "base J" en cinetoplasidos.
Dañar
El ADN puede ser dañado por muchos tipos de mutágenos, que cambian la secuencia de ADN. Los mutágenos incluyen agentes oxidantes, agentes alquilantes y también radiación electromagnética de alta energía, como la luz ultravioleta y los rayos X. El tipo de daño al ADN producido depende del tipo de mutágeno. Por ejemplo, la luz ultravioleta puede dañar el ADN produciendo dímeros de timina, que son enlaces cruzados entre bases de pirimidina. Por otro lado, oxidantes tales como radicales libres o peróxido de hidrógeno producen múltiples formas de daño, incluyendo modificaciones de bases, particularmente de guanosina, y roturas de doble cadena. Una célula humana típica contiene aproximadamente 150,000 bases que han sufrido daño oxidativo. De estas lesiones oxidativas, las más peligrosas son las roturas de doble cadena, ya que son difíciles de reparar y pueden producir mutaciones puntuales, inserciones, deleciones de la secuencia de ADN, y translocaciones cromosómicas. Estas mutaciones pueden causar cáncer. Debido a los límites inherentes en los mecanismos de reparación del ADN, si los humanos vivieran lo suficiente, eventualmente todos desarrollarían cáncer. Los daños en el ADN que ocurren naturalmente, debido a los procesos celulares normales que producen especies de oxígeno reactivas, las actividades hidrolíticas del agua celular, etc., también ocurren con frecuencia. Aunque la mayoría de estos daños se reparan, en cualquier celda puede quedar algún daño en el ADN a pesar de la acción de los procesos de reparación. Estos daños restantes en el ADN se acumulan con la edad en los tejidos postmitóticos de los mamíferos. Esta acumulación parece ser una importante causa subyacente del envejecimiento. Los daños en el ADN que ocurren naturalmente, debido a los procesos celulares normales que producen especies de oxígeno reactivas, las actividades hidrolíticas del agua celular, etc., también ocurren con frecuencia. Aunque la mayoría de estos daños se reparan, en cualquier celda puede quedar algún daño en el ADN a pesar de la acción de los procesos de reparación. Estos daños restantes en el ADN se acumulan con la edad en los tejidos postmitóticos de los mamíferos. Esta acumulación parece ser una importante causa subyacente del envejecimiento. Los daños en el ADN que ocurren naturalmente, debido a los procesos celulares normales que producen especies de oxígeno reactivas, las actividades hidrolíticas del agua celular, etc., también ocurren con frecuencia. Aunque la mayoría de estos daños se reparan, en cualquier celda puede quedar algún daño en el ADN a pesar de la acción de los procesos de reparación. Estos daños restantes en el ADN se acumulan con la edad en los tejidos postmitóticos de los mamíferos. Esta acumulación parece ser una importante causa subyacente del envejecimiento.
Muchos mutágenos encajan en el espacio entre dos pares de bases adyacentes, esto se llama intercalación . La mayoría de los intercaladores son moléculas aromáticas y planas; ejemplos incluyen bromuro de etidio, acridinas, daunomicina y doxorrubicina. Para que un intercalador encaje entre los pares de bases, las bases deben separarse, distorsionando las cadenas de ADN al desenrollar la doble hélice. Esto inhibe tanto la transcripción como la replicación del ADN, causando toxicidad y mutaciones. Como resultado, los intercaladores de ADN pueden ser carcinógenos y, en el caso de la talidomida, un teratógeno. Otros como benzo [ a] pireno diol epóxido y aflatoxina forman aductos de ADN que inducen errores en la replicación. Sin embargo, debido a su capacidad para inhibir la transcripción y la replicación del ADN, otras toxinas similares también se usan en la quimioterapia para inhibir el crecimiento rápido de las células cancerosas.
Funciones biológicas

El ADN generalmente ocurre como cromosomas lineales en eucariotas y cromosomas circulares en procariotas. El conjunto de cromosomas en una célula constituye su genoma; el genoma humano tiene aproximadamente 3 mil millones de pares de bases de ADN dispuestos en 46 cromosomas. La información transportada por el ADN se mantiene en la secuencia de fragmentos de ADN llamados genes. La transmisión de la información genética en los genes se logra mediante el emparejamiento de bases complementarias. Por ejemplo, en la transcripción, cuando una célula usa la información en un gen, la secuencia de ADN se copia en una secuencia de ARN complementario a través de la atracción entre el ADN y los nucleótidos de ARN correctos. Por lo general, esta copia de ARN se usa para hacer una secuencia de proteínas correspondiente en un proceso llamado traducción, que depende de la misma interacción entre los nucleótidos de ARN. De manera alternativa, una célula puede simplemente copiar su información genética en un proceso llamado replicación de ADN. Los detalles de estas funciones están cubiertos en otros artículos; aquí la atención se centra en las interacciones entre el ADN y otras moléculas que median en la función del genoma.
Genes y genomas
El ADN genómico se empaqueta de forma ajustada y ordenada en el proceso llamado condensación de ADN, para adaptarse a los pequeños volúmenes disponibles de la célula. En eucariotas, el ADN se encuentra en el núcleo de la célula, con pequeñas cantidades en las mitocondrias y los cloroplastos. En procariotas, el ADN se mantiene dentro de un cuerpo de forma irregular en el citoplasma llamado nucleoide. La información genética en un genoma se mantiene dentro de los genes, y el conjunto completo de esta información en un organismo se llama su genotipo. Un gen es una unidad de herencia y es una región de ADN que influye en una característica particular de un organismo. Los genes contienen un marco de lectura abierto que se puede transcribir, y secuencias reguladoras tales como promotores y potenciadores, que controlan la transcripción del marco de lectura abierto.
En muchas especies, solo una pequeña fracción de la secuencia total del genoma codifica proteína. Por ejemplo, solo alrededor del 1,5% del genoma humano consiste en exones que codifican proteínas, con más del 50% de ADN humano que consiste en secuencias repetitivas no codificantes. Las razones de la presencia de tanto ADN no codificante en los genomas eucariotas y las diferencias extraordinarias en el tamaño del genoma, o valor C , entre las especies, representan un rompecabezas de larga data conocido como el "enigma del valor C". Sin embargo, algunas secuencias de ADN que no codifican la proteína todavía pueden codificar moléculas de ARN funcionales no codificantes, que están implicadas en la regulación de la expresión génica.

Algunas secuencias de ADN no codificantes desempeñan papeles estructurales en los cromosomas. Los telómeros y los centrómeros suelen contener pocos genes, pero son importantes para la función y la estabilidad de los cromosomas. Una forma abundante de ADN no codificante en humanos son los pseudogenes, que son copias de genes que se han desactivado por mutación. Estas secuencias son generalmente solo fósiles moleculares, aunque ocasionalmente pueden servir como material genético crudo para la creación de nuevos genes a través del proceso de duplicación y divergencia de genes.
Transcripción y traducción
Un gen es una secuencia de ADN que contiene información genética y puede influir en el fenotipo de un organismo. Dentro de un gen, la secuencia de bases a lo largo de una cadena de ADN define una secuencia de ARN mensajero, que luego define una o más secuencias de proteína. La relación entre las secuencias de nucleótidos de los genes y las secuencias de aminoácidos de las proteínas está determinada por las reglas de la traducción, conocidas colectivamente como el código genético. El código genético consiste en "palabras" de tres letras llamadas codones formados a partir de una secuencia de tres nucleótidos (por ejemplo, ACT, CAG, TTT).
En la transcripción, los codones de un gen se copian en el ARN mensajero mediante la ARN polimerasa. Esta copia de ARN es decodificada por un ribosoma que lee la secuencia de ARN emparejando por bases el ARN mensajero para transferir ARN, que lleva aminoácidos. Como hay 4 bases en combinaciones de 3 letras, hay 64 posibles codones (4 combinaciones). Estos codifican los veinte aminoácidos estándar, dando a la mayoría de los aminoácidos más de un codón posible. También hay tres codones 'stop' o 'sinsentido' que significan el final de la región de codificación; estos son los codones TAA, TGA y TAG.
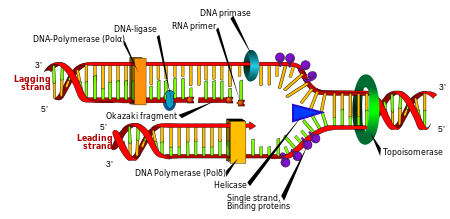
Replicación
La división celular es esencial para que un organismo crezca, pero cuando una célula se divide, debe replicar el ADN en su genoma para que las dos células hijas tengan la misma información genética que la de sus padres. La estructura bicatenaria del ADN proporciona un mecanismo simple para la replicación del ADN. Aquí, las dos cadenas se separan y luego la secuencia complementaria de ADN de cada cadena se recrea mediante una enzima llamada ADN polimerasa. Esta enzima forma la cadena complementaria al encontrar la base correcta a través del emparejamiento de bases complementarias y unirla a la cadena original. Como las ADN polimerasas solo pueden extender una cadena de ADN en una dirección de 5 'a 3', se usan diferentes mecanismos para copiar las cadenas antiparalelas de la doble hélice. De esta manera, la base en el viejo capítulo dicta qué base aparece en el nuevo capítulo,
Ácidos nucleicos extracelulares
El ADN extracelular desnudo (eADN), la mayoría liberado por la muerte celular, es casi omnipresente en el medio ambiente. Su concentración en el suelo puede ser tan alta como 2 μg / L, y su concentración en ambientes acuáticos naturales puede ser tan alta como 88 μg / L. Se han propuesto varias funciones posibles para eDNA: puede estar involucrado en la transferencia horizontal de genes; puede proporcionar nutrientes; y puede actuar como un tampón para reclutar o valorar iones o antibióticos. El ADN extracelular actúa como un componente funcional de la matriz extracelular en las biopelículas de varias especies bacterianas. Puede actuar como un factor de reconocimiento para regular el acoplamiento y la dispersión de tipos celulares específicos en la biopelícula; puede contribuir a la formación de biopelículas; y puede contribuir a la resistencia física y resistencia del biofilm al estrés biológico.
El ADN fetal libre de células se encuentra en la sangre de la madre y puede secuenciarse para determinar una gran cantidad de información sobre el desarrollo del feto.
Interacciones con proteínas
Todas las funciones del ADN dependen de las interacciones con las proteínas. Estas interacciones de proteínas pueden ser inespecíficas, o la proteína puede unirse específicamente a una sola secuencia de ADN. Las enzimas también pueden unirse al ADN y, de éstas, las polimerasas que copian la secuencia de bases de ADN en la transcripción y la replicación del ADN son particularmente importantes.
Proteínas de unión al ADN
 |
Interacción del ADN (en naranja) con las histonas (en azul). Los aminoácidos básicos de estas proteínas se unen a los grupos ácidos de fosfato en el ADN.
Las proteínas estructurales que se unen al ADN son ejemplos bien entendidos de interacciones ADN-proteína no específicas. Dentro de los cromosomas, el ADN se mantiene en complejos con proteínas estructurales. Estas proteínas organizan el ADN en una estructura compacta llamada cromatina. En eucariotas, esta estructura involucra el ADN que se une a un complejo de pequeñas proteínas básicas llamadas histonas, mientras que en procariotas se usan múltiples tipos de proteínas. Las histonas forman un complejo en forma de disco llamado nucleosoma, que contiene dos vueltas completas de ADN bicatenario envuelto alrededor de su superficie. Estas interacciones no específicas se forman a través de residuos básicos en las histonas, haciendo enlaces iónicos a la cadena principal ácida de azúcar y fosfato del ADN, y por lo tanto son en gran medida independientes de la secuencia de bases. Las modificaciones químicas de estos residuos de aminoácidos básicos incluyen metilación, fosforilación y acetilación. Estos cambios químicos alteran la fuerza de la interacción entre el ADN y las histonas, lo que hace que el ADN sea más o menos accesible para los factores de transcripción y el cambio de la tasa de transcripción. Otras proteínas de unión al ADN no específicas en la cromatina incluyen las proteínas del grupo de alta movilidad, que se unen al ADN doblado o distorsionado. Estas proteínas son importantes para doblar matrices de nucleosomas y organizarlas en las estructuras más grandes que componen los cromosomas.
Un grupo distinto de proteínas de unión a ADN son las proteínas de unión al ADN que se unen específicamente al ADN monocatenario. En humanos, la proteína de replicación A es el miembro mejor entendido de esta familia y se usa en procesos en los que se separa la doble hélice, incluida la replicación del ADN, la recombinación y la reparación del ADN. Estas proteínas de unión parecen estabilizar el ADN monocatenario y protegerlo de formar bucles del tallo o ser degradado por nucleasas.

Por el contrario, otras proteínas han evolucionado para unirse a secuencias de ADN particulares. Los más estudiados de estos son los diversos factores de transcripción, que son proteínas que regulan la transcripción. Cada factor de transcripción se une a un conjunto particular de secuencias de ADN y activa o inhibe la transcripción de genes que tienen estas secuencias cerca de sus promotores. Los factores de transcripción lo hacen de dos maneras. En primer lugar, pueden unirse a la ARN polimerasa responsable de la transcripción, ya sea directamente o a través de otras proteínas mediadoras; esto ubica la polimerasa en el promotor y le permite comenzar la transcripción. Alternativamente, los factores de transcripción pueden unirse a las enzimas que modifican las histonas en el promotor. Esto cambia la accesibilidad de la plantilla de ADN a la polimerasa.
Como estos objetivos de ADN pueden ocurrir en todo el genoma de un organismo, los cambios en la actividad de un tipo de factor de transcripción pueden afectar a miles de genes. Consecuentemente, estas proteínas suelen ser los objetivos de los procesos de transducción de señales que controlan las respuestas a los cambios ambientales o la diferenciación celular y el desarrollo . La especificidad de las interacciones de estos factores de transcripción con el ADN provienen de las proteínas que hacen contactos múltiples con los bordes de las bases de ADN, lo que les permite "leer" la secuencia de ADN. La mayoría de estas interacciones base se realizan en el surco mayor, donde las bases son más accesibles.

Enzimas que modifican el ADN
Nucleasas y ligasas
Las nucleasas son enzimas que cortan cadenas de ADN al catalizar la hidrólisis de los enlaces fosfodiéster. Las nucleasas que hidrolizan los nucleótidos de los extremos de las cadenas de ADN se denominan exonucleasas, mientras que las endonucleasas se cortan dentro de las cadenas. Las nucleasas utilizadas con mayor frecuencia en biología molecular son las endonucleasas de restricción, que cortan el ADN en secuencias específicas. Por ejemplo, la enzima EcoRV mostrada a la izquierda reconoce la secuencia de 6 bases 5'-GATATC-3 'y realiza un corte en la línea horizontal. En la naturaleza, estas enzimas protegen a las bacterias contra la infección del fago al digerir el ADN del fago cuando ingresa a la célula bacteriana, actuando como parte del sistema de modificación de restricción. En tecnología, estas nucleasas específicas de secuencia se usan en la clonación molecular y en la identificación del ADN.
Las enzimas llamadas ADN ligasas pueden unirse a cadenas de ADN cortadas o rotas. Las ligasas son particularmente importantes en la replicación del ADN filamento de revestimiento, ya que se unen los segmentos cortos de ADN producido en la replicación tenedor en una copia completa de la plantilla de ADN. También se usan en reparación de ADN y recombinación genética.
Topoisomerasas y helicasas
Las topoisomerasas son enzimas con actividad nucleasa y ligasa. Estas proteínas cambian la cantidad de superenrrollamiento en el ADN. Algunas de estas enzimas funcionan cortando la hélice del ADN y permitiendo que una sección gire, reduciendo así su nivel de superenrrollamiento; la enzima luego sella la ruptura del ADN. Otros tipos de estas enzimas son capaces de cortar una hélice de ADN y luego pasar una segunda cadena de ADN a través de esta ruptura, antes de volver a unirse a la hélice. Las topoisomerasas son necesarias para muchos procesos que implican ADN, como la replicación y la transcripción del ADN.
Helicases son proteínas que son un tipo de motor molecular. Utilizan la energía química en los trifosfatos de nucleósidos, predominantemente trifosfato de adenosina (ATP), para romper los enlaces de hidrógeno entre las bases y desenrollar la doble hélice del ADN en cadenas simples. Estas enzimas son esenciales para la mayoría de los procesos donde las enzimas necesitan acceder a las bases de ADN.
Polimerasas
Las polimerasas son enzimas que sintetizan cadenas de polinucleótidos a partir de trifosfatos de nucleósidos. La secuencia de sus productos se crea en base a cadenas de polinucleótidos existentes, que se llaman plantillas . Estas enzimas funcionan añadiendo repetidamente un nucleótido al grupo hidroxilo 3 'al final de la cadena de polinucleótidos en crecimiento. Como consecuencia, todas las polimerasas funcionan en una dirección de 5 'a 3'. En el sitio activo de estas enzimas, el nucleósido trifosfato entrante se empareja con la plantilla: esto permite que las polimerasas sinteticen con precisión la cadena complementaria de su molde. Las polimerasas se clasifican según el tipo de plantilla que utilizan.
En la replicación del ADN, las ADN polimerasas dependientes de ADN hacen copias de cadenas de polinucleótidos de ADN. Para preservar la información biológica, es esencial que la secuencia de bases en cada copia sea precisamente complementaria a la secuencia de bases en la cadena de plantilla. Muchas ADN polimerasas tienen una actividad de corrección de pruebas. Aquí, la polimerasa reconoce los errores ocasionales en la reacción de síntesis por la falta de emparejamiento de bases entre los nucleótidos mal emparejados. Si se detecta una falta de coincidencia, se activa una actividad de exonucleasa de 3 'a 5' y se elimina la base incorrecta. En la mayoría de los organismos, las ADN polimerasas funcionan en un gran complejo llamado replisoma que contiene múltiples subunidades accesorias, como la abrazadera de ADN o las helicasas.
Las ADN polimerasas dependientes de ARN son una clase especializada de polimerasas que copian la secuencia de una cadena de ARN en ADN. Incluyen la transcriptasa inversa, que es una enzima viral involucrada en la infección de las células por retrovirus, y la telomerasa, que es necesaria para la replicación de los telómeros. Por ejemplo, la transcriptasa inversa del VIH es una enzima para la replicación del virus del SIDA. La telomerasa es una polimerasa inusual porque contiene su propia plantilla de ARN como parte de su estructura. Sintetiza los telómeros en los extremos de los cromosomas. Los telómeros evitan la fusión de los extremos de los cromosomas vecinos y protegen los extremos cromosómicos del daño.
La transcripción se lleva a cabo mediante una ARN polimerasa dependiente de ADN que copia la secuencia de una cadena de ADN en ARN. Para comenzar a transcribir un gen, la ARN polimerasa se une a una secuencia de ADN llamada promotor y separa las cadenas de ADN. Luego copia la secuencia del gen en una transcripción de ARN mensajero hasta que alcanza una región de ADN llamada terminador, donde se detiene y se separa del ADN. Al igual que con las ADN polimerasas humanas dependientes del ADN, la ARN polimerasa II, la enzima que transcribe la mayoría de los genes en el genoma humano, opera como parte de un gran complejo de proteínas con múltiples subunidades reguladoras y accesorias.
Recombinación genética
 |
 |
Estructura de la unión Hollidayintermediate en recombinación genética. Las cuatro cadenas de ADN separadas son de color rojo, azul, verde y amarillo.

Una hélice de ADN generalmente no interactúa con otros segmentos de ADN, y en las células humanas, los diferentes cromosomas incluso ocupan áreas separadas en el núcleo llamadas "territorios cromosómicos". Esta separación física de diferentes cromosomas es importante para la capacidad del ADN para funcionar como un repositorio estable de información, ya que uno de los pocos momentos en que los cromosomas interactúan es en el cruce cromosómico, que ocurre durante la reproducción sexual, cuando ocurre la recombinación genética. Crossover cromosómico es cuando dos hélices de ADN se rompen, cambian una sección y luego vuelven a unirse.
La recombinación permite a los cromosomas intercambiar información genética y producir nuevas combinaciones de genes, lo que aumenta la eficiencia de la selección natural y puede ser importante en la rápida evolución de nuevas proteínas. La recombinación genética también puede estar involucrada en la reparación del ADN, particularmente en la respuesta de la célula a roturas de doble cadena.
La forma más común de cruce cromosómico es la recombinación homóloga, donde los dos cromosomas implicados comparten secuencias muy similares. La recombinación no homóloga puede ser perjudicial para las células, ya que puede producir translocaciones cromosómicas y anomalías genéticas. La reacción de recombinación es catalizada por enzimas conocidas como recombinasas, como RAD51. El primer paso en la recombinación es una ruptura bicatenaria causada por una endonucleasa o daño al ADN. Una serie de etapas catalizadas en parte por la recombinasa conduce luego a la unión de las dos hélices por al menos una unión de Holliday, en la que un segmento de una sola cadena en cada hélice se hibrida con la cadena complementaria en la otra hélice. La unión de Holliday es una estructura de unión tetraédrica que se puede mover a lo largo del par de cromosomas, intercambiando un filamento por otro. La reacción de recombinación se detiene después mediante la escisión de la unión y la ligación de nuevo del ADN liberado. Solo las cadenas de polaridad similar intercambian ADN durante la recombinación. Hay dos tipos de escote: escote este-oeste y escote norte-sur. La división norte-sur mella ambas hebras de ADN, mientras que la hendidura este-oeste tiene una hebra de ADN intacta. La formación de una unión de Holliday durante la recombinación hace posible la diversidad genética, el intercambio de genes en los cromosomas y la expresión de genomas virales de tipo salvaje. mientras que la división este-oeste tiene una hebra de ADN intacta. La formación de una unión de Holliday durante la recombinación hace posible la diversidad genética, el intercambio de genes en los cromosomas y la expresión de genomas virales de tipo salvaje. mientras que la división este-oeste tiene una hebra de ADN intacta. La formación de una unión de Holliday durante la recombinación hace posible la diversidad genética, el intercambio de genes en los cromosomas y la expresión de genomas virales de tipo salvaje.
Evolución
El ADN contiene la información genética que permite que todos los seres vivos modernos funcionen, crezcan y se reproduzcan. Sin embargo, no está claro por cuánto tiempo en el historial de vida de 4 mil millones de años el ADN ha llevado a cabo esta función, ya que se ha propuesto que las primeras formas de vida pueden haber utilizado el ARN como material genético. El ARN puede haber actuado como la parte central del metabolismo celular temprano, ya que puede transmitir información genética y llevar a cabo la catálisis como parte de las ribozimas. Este antiguo mundo de ARN donde el ácido nucleico se habría usado tanto para la catálisis como para la genética, puede haber influido en la evolución del código genético actual basado en cuatro bases de nucleótidos. Esto ocurriría, dado que el número de bases diferentes en dicho organismo es una compensación entre un pequeño número de bases que aumenta la precisión de la replicación y un gran número de bases que aumentan la eficacia catalítica de las ribozimas. Sin embargo, no hay evidencia directa de sistemas genéticos antiguos, ya que la recuperación de ADN de la mayoría de los fósiles es imposible porque el ADN sobrevive en el medio ambiente durante menos de un millón de años y se degrada lentamente en fragmentos cortos en solución. Se han realizado reclamos por ADN más antiguo, sobre todo un informe del aislamiento de una bacteria viable de un cristal de sal de hace 250 millones de años, pero estas afirmaciones son controvertidas. y lentamente se degrada en fragmentos cortos en solución. Se han realizado reclamos por ADN más antiguo, sobre todo un informe del aislamiento de una bacteria viable de un cristal de sal de hace 250 millones de años, pero estas afirmaciones son controvertidas. y lentamente se degrada en fragmentos cortos en solución. Se han realizado reclamos por ADN más antiguo, sobre todo un informe del aislamiento de una bacteria viable de un cristal de sal de hace 250 millones de años, pero estas afirmaciones son controvertidas.
Los bloques de construcción de ADN (adenina, guanina y moléculas orgánicas relacionadas) pueden haberse formado extraterrestremente en el espacio exterior. También se han formado compuestos de vida orgánicos complejos de ADN y ARN, que incluyen uracilo, citosina y timina, en condiciones similares a las que se encuentran en el espacio exterior, utilizando productos químicos de partida, como la pirimidina, que se encuentra en los meteoritos. La pirimidina, como los hidrocarburos aromáticos policíclicos (HAP), la sustancia química más rica en carbono encontrada en el universo, puede haberse formado en gigantes rojos o en polvo cósmico interestelar y nubes de gas.
Usos en tecnología
Ingeniería genética
Se han desarrollado métodos para purificar el ADN de organismos, como la extracción con fenol-cloroformo, y para manipularlo en el laboratorio, como digestiones de restricción y la reacción en cadena de la polimerasa. La biología moderna y la bioquímica hacen un uso intensivo de estas técnicas en la tecnología del ADN recombinante. El ADN recombinante es una secuencia de ADN artificial que se ha ensamblado a partir de otras secuencias de ADN. Se pueden transformar en organismos en forma de plásmidos o en el formato apropiado, utilizando un vector viral. Los organismos modificados genéticamente producidos pueden usarse para producir productos tales como proteínas recombinantes, usados en investigaciones médicas o cultivados en la agricultura.
Perfiles de ADN
Los científicos forenses pueden usar ADN en la sangre, el semen, la piel, la saliva o el cabello que se encuentran en la escena del crimen para identificar un ADN correspondiente de un individuo, como un perpetrador. Este proceso se denomina formalmente perfil de ADN, pero también se lo puede llamar "huella digital genética". En el perfil de ADN, se comparan las longitudes de las secciones variables de ADN repetitivo, como las repeticiones cortas en tándem y los minisatélites, entre las personas. Este método es usualmente una técnica extremadamente confiable para identificar un ADN coincidente. Sin embargo, la identificación puede ser complicada si la escena está contaminada con ADN de varias personas. La creación de perfiles de ADN fue desarrollada en 1984 por el genetista británico Sir Alec Jeffreys, y se usó por primera vez en la ciencia forense para condenar a Colin Pitchfork en el caso de asesinatos de Enderby en 1988.
El desarrollo de la ciencia forense y la capacidad de obtener ahora coincidencias genéticas en muestras diminutas de sangre, piel, saliva o cabello ha llevado a reexaminar muchos casos. Ahora se puede descubrir evidencia que era científicamente imposible en el momento del examen original. Combinado con la eliminación de la ley de doble riesgo en algunos lugares, esto puede permitir que los casos se vuelvan a abrir donde los ensayos anteriores no han podido producir pruebas suficientes para convencer a un jurado. A las personas acusadas de delitos graves se les puede solicitar que proporcionen una muestra de ADN para fines de comparación. La defensa más obvia para las coincidencias de ADN obtenidas de manera forense es afirmar que se ha producido una contaminación cruzada de la evidencia. Esto ha resultado en procedimientos de manejo estrictos y meticulosos con nuevos casos de delitos graves.
El perfil de ADN también se utiliza en pruebas de paternidad de ADN para determinar si alguien es el padre biológico o abuelo de un niño con probabilidad de paternidad es típicamente 99.99% cuando el supuesto padre está relacionado biológicamente con el niño. Los métodos normales de secuenciación del ADN ocurren después del nacimiento, pero existen nuevos métodos para evaluar la paternidad mientras la madre todavía está embarazada.
Enzimas de ADN o ADN catalítico
Las desoxirribozimas, también llamadas ADNzimas o ADN catalítico, se descubrieron por primera vez en 1994. Son en su mayoría secuencias de ADN monocatenarias aisladas de un gran conjunto de secuencias aleatorias de ADN mediante un enfoque combinatorio denominado selección in vitro o evolución sistemática de ligandos por enriquecimiento exponencial (SELEX) . Las ADNzimas catalizan una variedad de reacciones químicas que incluyen escisión ARN-ADN, ligadura ARN-ADN, fosforilación-desfosforilación de aminoácidos, formación de enlaces carbono-carbono, etc. Las ADNzimas pueden aumentar la tasa catalítica de reacciones químicas hasta 100,000,000,000 veces sobre la reacción no catalizada. La clase más ampliamente estudiada de ADNzimas son los tipos de escisión de ARN que se han usado para detectar diferentes iones metálicos y diseñar agentes terapéuticos. Se han informado varias ADNzimas específicas de metales, incluida la ADNzima GR-5 (específica del plomo), las ADNzimas CA1-3 (específicas de cobre), la ADNzima 39E (específica de uranilo) y la ADNzima NaA43 (específica de sodio). La ADNzima NaA43, que se informa que es más de 10.000 veces selectiva para el sodio en comparación con otros iones metálicos, se usó para hacer un sensor de sodio en tiempo real en las células vivas.
Bioinformática
La bioinformática implica el desarrollo de técnicas para almacenar, extraer datos, buscar y manipular datos biológicos, incluidos datos de secuencias de ácidos nucleicos de ADN. Esto ha llevado a avances ampliamente aplicados en ciencias de la computación, especialmente algoritmos de búsqueda de cadenas, aprendizaje automático y teoría de bases de datos. La búsqueda de cadenas o algoritmos de coincidencia, que encuentran una ocurrencia de una secuencia de letras dentro de una secuencia más grande de letras, se desarrollaron para buscar secuencias específicas de nucleótidos. La secuencia de ADN puede alinearse con otras secuencias de ADN para identificar secuencias homólogas y localizar las mutaciones específicas que las hacen distintas. Estas técnicas, especialmente la alineación de secuencias múltiples, se utilizan en el estudio de las relaciones filogenéticas y la función proteica. Conjuntos de datos que representan las secuencias de ADN de genomas completos como los producidos por el Proyecto del Genoma Humano, son difíciles de usar sin las anotaciones que identifican las ubicaciones de los genes y los elementos reguladores en cada cromosoma. Las regiones de secuencia de ADN que tienen los patrones característicos asociados a genes codificantes de proteína o ARN pueden identificarse mediante algoritmos de búsqueda de genes, que permiten a los investigadores predecir la presencia de productos genéticos particulares y sus posibles funciones en un organismo incluso antes de que se hayan aislado. experimentalmente. También se pueden comparar genomas completos, que pueden arrojar luz sobre la historia evolutiva de un organismo particular y permitir el examen de eventos evolutivos complejos. Las regiones de secuencia de ADN que tienen los patrones característicos asociados a genes codificantes de proteína o ARN pueden identificarse mediante algoritmos de búsqueda de genes, que permiten a los investigadores predecir la presencia de productos genéticos particulares y sus posibles funciones en un organismo incluso antes de que se hayan aislado. experimentalmente. También se pueden comparar genomas completos, que pueden arrojar luz sobre la historia evolutiva de un organismo particular y permitir el examen de eventos evolutivos complejos. Las regiones de secuencia de ADN que tienen los patrones característicos asociados a genes codificantes de proteína o ARN pueden identificarse mediante algoritmos de búsqueda de genes, que permiten a los investigadores predecir la presencia de productos genéticos particulares y sus posibles funciones en un organismo incluso antes de que se hayan aislado. experimentalmente. También se pueden comparar genomas completos, que pueden arrojar luz sobre la historia evolutiva de un organismo particular y permitir el examen de eventos evolutivos complejos.
Nanotecnología del ADN

La nanotecnología del ADN utiliza las propiedades únicas de reconocimiento molecular del ADN y otros ácidos nucleicos para crear complejos de ADN ramificado autoensamblables con propiedades útiles. Por lo tanto, el ADN se usa como material estructural en lugar de como portador de información biológica. Esto ha llevado a la creación de celosías periódicas bidimensionales (basadas en mosaicos y utilizando el método de origami de ADN ) y estructuras tridimensionales en forma de poliedros. También se han demostrado dispositivos nanomecánicos y autoensamblaje algorítmico, y estas estructuras de ADN se han utilizado para moldear la disposición de otras moléculas, como las nanopartículas de oro y las proteínas de estreptavidina.
Historia y antropología
Debido a que el ADN recolecta mutaciones a lo largo del tiempo, que luego son heredadas, contiene información histórica y, al comparar secuencias de ADN, los genetistas pueden inferir la historia evolutiva de los organismos, su filogenia. Este campo de la filogenética es una poderosa herramienta en biología evolutiva. Si se comparan secuencias de ADN dentro de una especie, los genetistas de poblaciones pueden aprender la historia de poblaciones particulares. Esto se puede utilizar en estudios que van desde la genética ecológica hasta la antropología.
Almacenamiento de informacion
En un artículo publicado en Nature en enero de 2013, científicos del Instituto Europeo de Bioinformática y Agilent Technologies propusieron un mecanismo para utilizar la capacidad del ADN para codificar la información como un medio de almacenamiento de datos digitales. El grupo pudo codificar 739 kilobytes de datos en código de ADN, sintetizar el ADN real, luego secuenciar el ADN y decodificar la información a su forma original, con una precisión del 100%. La información codificada consistía en archivos de texto y archivos de audio. Un experimento previo fue publicado en agosto de 2012. Fue realizado por investigadores de la Universidad de Harvard, donde el texto de un libro de 54,000 palabras estaba codificado en ADN.
Además, en las células vivas, las enzimas pueden activar el almacenamiento. Los dominios de proteínas de luz controlada fusionados a enzimas de procesamiento de ADN son adecuados para esa tarea in vitro . Las exonucleasas fluorescentes pueden transmitir la salida de acuerdo con el nucleótido que han leído.
Historia

El ADN fue aislado por primera vez por el médico suizo Friedrich Miescher que, en 1869, descubrió una sustancia microscópica en el pus de los vendajes quirúrgicos desechados. Como residía en el núcleo de las células, lo llamó "nucleina". En 1878, Albrecht Kossel aisló el componente no proteico de "nucleina", ácido nucleico, y posteriormente aisló sus cinco nucleobases primarios.
En 1909, Phoebus Levene identificó la unidad de nucleótidos de base, azúcar y fosfato del ARN (entonces llamada "ácido nucleico de levadura"). En 1929, Levene identificó el azúcar desoxirribosa en el "ácido nucleico del timo" (ADN). Levene sugirió que el ADN consistía en una cadena de cuatro unidades de nucleótidos unidas mediante los grupos fosfato ("hipótesis de tetranucleótidos"). Levene pensó que la cadena era corta y las bases se repitieron en un orden fijo. En 1927, Nikolai Koltsov propuso que los rasgos heredados se heredarían a través de una "molécula hereditaria gigante" compuesta de "dos hebras de espejo que se replicarían de forma semi-conservativa utilizando cada hebra como plantilla". En 1928, Frederick Griffith en su experimento descubrió los rasgos de la forma "suave" de Pneumococcus podría transferirse a la forma "rugosa" de la misma bacteria mezclando bacterias muertas "lisas" con la forma viva "áspera". Este sistema proporcionó la primera sugerencia clara de que el ADN transporta información genética.
En 1933, mientras estudiaba huevos de erizo de mar virgen, Jean Brachet sugirió que el ADN se encuentra en el núcleo de la célula y que el ARN está presente exclusivamente en el citoplasma. En ese momento, se pensaba que el "ácido nucleico de la levadura" (ARN) solo se producía en las plantas, mientras que el "ácido nucleico del timo" (ADN) solo en los animales. Se pensaba que este último era un tetrámero, con la función de amortiguar el pH celular.
En 1937, William Astbury produjo los primeros patrones de difracción de rayos X que demostraban que el ADN tenía una estructura regular.
En 1943, Oswald Avery, junto con sus compañeros de trabajo Colin MacLeod y Maclyn McCarty, identificaron el ADN como el principio transformador, apoyando la sugerencia de Griffith (experimento Avery-MacLeod-McCarty). El papel del ADN en la herencia se confirmó en 1952 cuando Alfred Hershey y Martha Chase en el experimento Hershey-Chase demostraron que el ADN es el material genético del fago T2.

A fines de 1951, Francis Crick comenzó a trabajar con James Watson en el Laboratorio Cavendish de la Universidad de Cambridge. En 1953, Watson y Crick sugirieron lo que ahora se acepta como el primer modelo correcto de doble hélice de la estructura del ADN en la revista Nature . Su modelo de ADN de doble hélice se basó en una imagen de difracción de rayos X (etiquetada como "Foto 51") tomada por Rosalind Franklin y Raymond Gosling en mayo de 1952, y la información de que las bases de ADN están emparejadas. El 28 de febrero de 1953, Crick interrumpió el almuerzo de los clientes en el pub The Eagle de Cambridge para anunciar que él y Watson habían "descubierto el secreto de la vida".
Meses antes, en febrero de 1953, Linus Pauling y Robert Corey propusieron un modelo para ácidos nucleicos que contenía tres cadenas entrelazadas, con los fosfatos cerca del eje y las bases en el exterior. La evidencia experimental que respalda el modelo de Watson y Crick se publicó en una serie de cinco artículos en el mismo número de Nature . De estos, el artículo de Franklin y Gosling fue la primera publicación de sus propios datos de difracción de rayos X y el método de análisis original que en parte respaldaban el modelo de Watson y Crick; este tema también contenía un artículo sobre la estructura del ADN de Maurice Wilkins y dos de sus colegas, cuyo análisis y patrones de rayos-X de ADN-B in vivo también respaldaban la presencia in vivo de las configuraciones de doble hélice de ADN propuestas por Crick y Watson para su modelo molecular de doble hélice de ADN en las dos páginas anteriores de Nature . En 1962, después de la muerte de Franklin, Watson, Crick y Wilkins recibieron conjuntamente el Premio Nobel de Fisiología o Medicina. Los Premios Nobel se otorgan solo a los destinatarios vivos. Continúa un debate sobre quién debe recibir crédito por el descubrimiento.
En una presentación influyente en 1957, Crick expuso el dogma central de la biología molecular, que predijo la relación entre el ADN, el ARN y las proteínas, y articuló la "hipótesis del adaptador". La confirmación final del mecanismo de replicación que implicaba la estructura de doble hélice siguió en 1958 a través del experimento Meselson-Stahl. El trabajo adicional de Crick y sus colegas mostró que el código genético se basaba en triples de bases no superpuestos, llamados codones, lo que permitía a Har Gobind Khorana, Robert W. Holley y Marshall Warren Nirenberg descifrar el código genético. Estos hallazgos representan el nacimiento de la biología molecular.