Gen
Definición
En biología, un gen es una secuencia de ADN o ARN que codifica una molécula que tiene una función. Durante la expresión génica, el ADN se copia primero en ARN. El ARN puede ser directamente funcional o ser la plantilla intermedia para una proteína que realiza una función. La transmisión de genes a la descendencia de un organismo es la base de la herencia de los rasgos fenotípicos. Estos genes componen diferentes secuencias de ADN llamadas genotipos. Los genotipos junto con los factores ambientales y de desarrollo determinan cuáles serán los fenotipos. La mayoría de los rasgos biológicos están bajo la influencia de polygenes (muchos genes diferentes) así como de interacciones gen-ambiente. Algunos rasgos genéticos son instantáneamente visibles, como el color de ojos o la cantidad de miembros, y otros no, como el tipo de sangre, el riesgo de enfermedades específicas o los miles de procesos bioquímicos básicos que constituyen la vida.Los genes pueden adquirir mutaciones en su secuencia, dando lugar a diferentes variantes, conocidas como alelos, en la población. Estos alelos codifican versiones ligeramente diferentes de una proteína, que causan diferentes características fenotípicas. El uso del término "tener un gen" (por ejemplo, "genes buenos", "gen del color del cabello") típicamente se refiere a que contiene un alelo diferente del mismo gen compartido. Los genes evolucionan debido a la selección natural o la supervivencia del alelo más apto.
El concepto de un gen continúa siendo refinado a medida que se descubren nuevos fenómenos. Por ejemplo, las regiones reguladoras de un gen pueden estar muy alejadas de sus regiones codificantes, y las regiones codificantes pueden dividirse en varios exones. Algunos virus almacenan su genoma en ARN en lugar de ADN y algunos productos genéticos son ARN funcionales no codificantes. Por lo tanto, una definición de trabajo amplia y moderna de un gen es cualquier locus discreto de secuencia genómica hereditaria que afecta los rasgos de un organismo al expresarse como un producto funcional o mediante la regulación de la expresión génica.
El término gen fue introducido por el botánico, fisiólogo y genetista danés Wilhelm Johannsen en 1905. Está inspirado en el griego antiguo: γόνος, gonos , que significa descendencia y procreación.
Historia

Descubrimiento de unidades heredadas discretas
La existencia de unidades heredables discretas fue sugerida por primera vez por Gregor Mendel (1822-1884). De 1857 a 1864, en Brno (República Checa), estudió los patrones de herencia en 8000 plantas de guisantes comestibles comunes, rastreando rasgos distintivos de padres a hijos. Él los describió matemáticamente como 2 combinaciones donde n es el número de características diferentes en los guisantes originales. Aunque no usó el término gen, explicó sus resultados en términos de unidades heredadas discretas que dan lugar a características físicas observables. Esta descripción prefigura la distinción de Wilhelm Johannsen entre el genotipo (el material genético de un organismo) y el fenotipo (los rasgos visibles de ese organismo). Mendel también fue el primero en demostrar el surtido independiente, la distinción entre rasgos dominantes y recesivos, la distinción entre un heterocigoto y homocigoto, y el fenómeno de la herencia discontinua.
Antes del trabajo de Mendel, la teoría dominante de la herencia era la herencia de mezcla, que sugería que cada padre aportaba fluidos para el proceso de fertilización y que los rasgos de los padres se mezclaban y mezclaban para producir la descendencia. Charles Darwin desarrolló una teoría de la herencia que denominó pangénesis, del griego pan ("todo, entero") y génesis ("nacimiento") / genos ("origen"). Darwin usó el término gemmule para describir partículas hipotéticas que se mezclarían durante la reproducción.
El trabajo de Mendel pasó desapercibido después de su primera publicación en 1866, pero fue redescubierto a finales del siglo XIX por Hugo de Vries, Carl Correns y Erich von Tschermak, quienes (según dicen haberlo hecho) llegaron a conclusiones similares en sus propias investigaciones. Específicamente, en 1889, Hugo de Vries publicó su libro Pangenesis intracelular , en el cual postuló que diferentes personajes tienen portadores hereditarios individuales y que la herencia de rasgos específicos en los organismos viene en partículas. De Vries llamó a estas unidades " pangenes " ( Pangens en alemán), después de la teoría de la pangénesis de 1868 de Darwin.
Dieciséis años después, en 1905, Wilhelm Johannsen introdujo el término "gen" y William Bateson el de "genética", mientras que Eduard Strasburger, entre otros, todavía utilizaba el término "pangene" para la unidad física y funcional fundamental de la herencia.
Descubrimiento de ADN
Los avances en la comprensión de los genes y la herencia continuaron a lo largo del siglo XX. Se demostró que el ácido desoxirribonucleico (ADN) es el depósito molecular de información genética mediante experimentos en los años 1940 a 1950. La estructura del ADN fue estudiada por Rosalind Franklin y Maurice Wilkins utilizando cristalografía de rayos X, lo que llevó a James D. Watson y Francis Crick a publicar un modelo de la molécula de ADN bicatenario cuyas bases de nucleótidos apareados indicaban una hipótesis convincente para el mecanismo de replicación genética
A principios de la década de 1950, la opinión predominante era que los genes en un cromosoma actuaban como entidades discretas, indivisibles por recombinación y dispuestos como cuentas en una cuerda. Los experimentos de Benzer utilizando mutantes defectuosos en la región rII del bacteriófago T4 (1955-1959) mostraron que los genes individuales tienen una estructura lineal simple y es probable que sean equivalentes a una sección lineal de ADN.
Colectivamente, este cuerpo de investigación estableció el dogma central de la biología molecular, que establece que las proteínas se traducen del ARN, que se transcribe a partir del ADN. Desde entonces, se ha demostrado que este dogma tiene excepciones, como la transcripción inversa en retrovirus. El estudio moderno de la genética a nivel de ADN se conoce como genética molecular.
En 1972, Walter Fiers y su equipo fueron los primeros en determinar la secuencia de un gen: el de la proteína de la cubierta del bacteriófago MS2. El posterior desarrollo de la secuenciación de ADN de terminación de cadena en 1977 por Frederick Sanger mejoró la eficiencia de la secuenciación y la convirtió en una herramienta de laboratorio de rutina. Se utilizó una versión automatizada del método de Sanger en las primeras fases del Proyecto del Genoma Humano.
Síntesis moderna y sus sucesores
Las teorías desarrolladas a principios del siglo XX para integrar la genética mendeliana con la evolución darwiniana se llaman síntesis moderna, un término introducido por Julian Huxley.
Los biólogos evolutivos han modificado posteriormente este concepto, como la visión de la evolución centrada en el gen de George C. Williams. Propuso un concepto evolutivo del gen como una unidad de selección natural con la definición: "aquello que segrega y recombina con frecuencia apreciable". En esta visión, el gen molecular transcribe como una unidad, y el gen evolutivo hereda como una unidad. Las ideas relacionadas que enfatizan la centralidad de los genes en la evolución fueron popularizadas por Richard Dawkins.
Bases moleculares
ADN
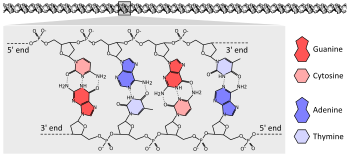
La gran mayoría de los organismos vivos codifican sus genes en largas cadenas de ADN (ácido desoxirribonucleico). El ADN consiste en una cadena compuesta por cuatro tipos de subunidades de nucleótidos, cada una compuesta por: un azúcar de cinco carbonos (2'-desoxirribosa), un grupo fosfato y una de las cuatro bases adenina, citosina, guanina y timina.
Dos cadenas de ADN giran una alrededor de la otra para formar una doble hélice de ADN con la cadena principal de azúcar fosfato girando en espiral hacia afuera, y las bases apuntando hacia adentro con la base de adenina que se combina con la timina y la guanina con la citosina. La especificidad del emparejamiento de bases ocurre porque la adenina y la timina se alinean para formar dos enlaces de hidrógeno, mientras que la citosina y la guanina forman tres enlaces de hidrógeno. Las dos hebras en una doble hélice deben, por lo tanto, ser complementarias, con su secuencia de bases que coincida de tal manera que las adeninas de una cadena estén emparejadas con las timinas de la otra cadena, y así sucesivamente.
Debido a la composición química de los residuos de pentosa de las bases, las cadenas de ADN tienen direccionalidad. Un extremo de un polímero de ADN contiene un grupo hidroxilo expuesto en la desoxirribosa; esto se conoce como el extremo 3 'de la molécula. El otro extremo contiene un grupo fosfato expuesto; este es el 5 'final. Las dos hebras de una doble hélice corren en direcciones opuestas. La síntesis de ácido nucleico, incluida la replicación y transcripción del ADN, se produce en la dirección 5 '→ 3', porque se añaden nuevos nucleótidos a través de una reacción de deshidratación que utiliza el hidroxilo 3 'expuesto como un nucleófilo.
La expresión de genes codificados en el ADN comienza transcribiendo el gen en ARN, un segundo tipo de ácido nucleico que es muy similar al ADN, pero cuyos monómeros contienen el azúcar ribosa más que la desoxirribosa. El ARN también contiene el uracilo de base en lugar de timina. Las moléculas de ARN son menos estables que el ADN y típicamente son monocatenarias. Los genes que codifican proteínas se componen de una serie de secuencias de tres nucleótidos llamadas codones, que sirven como las "palabras" en el "lenguaje" genético. El código genético especifica la correspondencia durante la traducción de proteínas entre codones y aminoácidos. El código genético es casi el mismo para todos los organismos conocidos.
Cromosomas
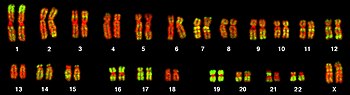
El complemento total de genes en un organismo o célula se conoce como su genoma, que puede almacenarse en uno o más cromosomas. Un cromosoma consiste en una sola hélice de ADN muy larga en la que se codifican miles de genes. La región del cromosoma en el que se encuentra un gen particular se llama su locus. Cada locus contiene un alelo de un gen; sin embargo, los miembros de una población pueden tener alelos diferentes en el locus, cada uno con una secuencia de gen ligeramente diferente.
La mayoría de los genes eucarióticos se almacenan en un conjunto de cromosomas lineales grandes. Los cromosomas se empaquetan dentro del núcleo en complejo con proteínas de almacenamiento llamadas histonas para formar una unidad llamada nucleosoma. El ADN empaquetado y condensado de esta manera se llama cromatina. La manera en que el ADN se almacena en las histonas, así como las modificaciones químicas de la misma histona, regulan si una región particular de ADN es accesible para la expresión génica. Además de los genes, los cromosomas eucarióticos contienen secuencias implicadas en garantizar que el ADN se copie sin degradación de las regiones terminales y se clasifique en células hijas durante la división celular: orígenes de replicación, telómeros y centrómero. Los orígenes de replicación son las regiones de secuencia donde se inicia la replicación del ADN para hacer dos copias del cromosoma. Los telómeros son largos tramos de secuencia repetitiva que cubren los extremos de los cromosomas lineales y evitan la degradación de las regiones codificadoras y reguladoras durante la replicación del ADN. La longitud de los telómeros disminuye cada vez que el genoma se replica y se ha implicado en el proceso de envejecimiento. El centrómero es necesario para unir las fibras del huso para separar las cromátidas hermanas en las células hijas durante la división celular.
Los procariotas (bacterias y arqueas) generalmente almacenan sus genomas en un único cromosoma circular grande. Del mismo modo, algunos orgánulos eucarióticos contienen un cromosoma circular remanente con un pequeño número de genes. Los procariotas a veces complementan su cromosoma con pequeños círculos adicionales de ADN llamados plásmidos, que generalmente codifican solo unos pocos genes y son transferibles entre individuos. Por ejemplo, los genes para la resistencia a los antibióticos normalmente están codificados en plásmidos bacterianos y pueden transmitirse entre células individuales, incluso las de diferentes especies, mediante transferencia génica horizontal.
Mientras que los cromosomas de los procariotas son relativamente genéticamente densos, los de los eucariotas a menudo contienen regiones de ADN que no cumplen ninguna función obvia. Los eucariotas unicelulares simples tienen cantidades relativamente pequeñas de dicho ADN, mientras que los genomas de organismos multicelulares complejos, incluidos los humanos, contienen una mayoría absoluta de ADN sin una función identificada. Este ADN a menudo se conoce como "ADN basura". Sin embargo, análisis más recientes sugieren que, aunque el ADN que codifica proteínas constituye apenas el 2% del genoma humano, aproximadamente el 80% de las bases en el genoma pueden expresarse, por lo que el término "ADN basura" puede ser un nombre inapropiado.
Estructura y función
Estructura
La estructura de un gen consiste en muchos elementos de los cuales la secuencia de codificación de proteína real a menudo es solo una pequeña parte. Estos incluyen regiones de ADN que no se transcriben, así como regiones no traducidas del ARN.
Flanqueando el marco de lectura abierto, los genes contienen una secuencia regulatoria que se requiere para su expresión. Primero, los genes requieren una secuencia promotora. El promotor es reconocido y unido por factores de transcripción y ARN polimerasa para iniciar la transcripción. El reconocimiento normalmente ocurre como una secuencia de consenso como la caja TATA. Un gen puede tener más de un promotor, dando como resultado ARN mensajeros (ARNm) que difieren en cuánto se extienden en el extremo 5 '. Los genes altamente transcritos tienen secuencias promotoras "fuertes" que forman fuertes asociaciones con factores de transcripción, iniciando así la transcripción a una velocidad elevada. Otros genes tienen promotores "débiles" que forman asociaciones débiles con factores de transcripción e inician la transcripción con menos frecuencia.
Además, los genes pueden tener regiones reguladoras de muchas kilobases aguas arriba o aguas abajo del marco de lectura abierto que alteran la expresión. Estos actúan uniéndose a factores de transcripción que luego hacen que el ADN se bucle de modo que la secuencia reguladora (y el factor de transcripción unido) se vuelva cercana al sitio de unión de la ARN polimerasa. Por ejemplo, los potenciadores aumentan la transcripción uniendo una proteína activadora que luego ayuda a reclutar la ARN polimerasa al promotor; por el contrario, los silenciadores se unen a las proteínas represoras y hacen que el ADN esté menos disponible para la ARN polimerasa.
El pre-ARNm transcrito contiene regiones no traducidas en ambos extremos que contienen un sitio de unión al ribosoma, terminador y codones de inicio y finalización. Además, la mayoría de los marcos de lectura abiertos eucarióticos contienen intrones no traducidos que se eliminan antes de que se traduzcan los exones. Las secuencias en los extremos de los intrones, dictan los sitios de empalme para generar el ARNm maduro final que codifica la proteína o producto de ARN.
Muchos genes procariotas se organizan en operones, con múltiples secuencias de codificación de proteínas que se transcriben como una unidad. Los genes en un operón se transcriben como un ARN mensajero continuo, denominado ARNm policistrónico. El término cistrón en este contexto es equivalente al gen. La transcripción del ARNm de un operón a menudo está controlada por un represor que puede ocurrir en un estado activo o inactivo dependiendo de la presencia de ciertos metabolitos específicos. Cuando está activo, el represor se une a una secuencia de ADN al comienzo del operón, llamada región operadora, y reprime la transcripción del operón; cuando el represor es inactivo, puede producirse la transcripción del operón (véase, por ejemplo, el operón Lac). Los productos de los genes del operón típicamente tienen funciones relacionadas y están involucrados en la misma red regulatoria.
Definiciones funcionales
Definir exactamente qué sección de una secuencia de ADN comprende un gen es difícil. Las regiones reguladoras de un gen como potenciadores no necesariamente tienen que estar cerca de la secuencia codificadora de la molécula lineal porque el ADN intermedio se puede bifurcar para llevar el gen y su región reguladora en la proximidad. Del mismo modo, los intrones de un gen pueden ser mucho más grandes que sus exones. Las regiones reguladoras pueden incluso estar en cromosomas completamente diferentes y operar en trans para permitir que las regiones reguladoras en un cromosoma entren en contacto con genes diana en otro cromosoma.
Los primeros trabajos en genética molecular sugirieron el concepto de que un gen produce una proteína. Este concepto (originalmente llamado la hipótesis del gen uno enzima) surgió de un influyente artículo de 1941 de George Beadle y Edward Tatum sobre experimentos con mutantes del hongo Neurospora crassa. Norman Horowitz, un antiguo colega en la investigación de Neurospora, recordó en 2004 que "estos experimentos fundaron la ciencia de lo que Beadle y Tatum llamaron genética bioquímica".. En realidad, demostraron ser la pistola de apertura en lo que se convirtió en genética molecular y todos los desarrollos que han seguido a partir de ahí. "El concepto de proteína de un solo gen se ha refinado desde el descubrimiento de genes que pueden codificar proteínas múltiples por corte y empalme alternativo las secuencias se dividen en una sección corta a través del genoma cuyos ARNm se concatenan por empalme trans.
A veces se utiliza una definición operacional amplia para abarcar la complejidad de estos diversos fenómenos, donde un gen se define como una unión de secuencias genómicas que codifican un conjunto coherente de productos funcionales potencialmente superpuestos. Esta definición clasifica los genes por sus productos funcionales (proteínas o ARN) en lugar de sus loci de ADN específicos, con elementos reguladores clasificados como regiones asociadas a genes .
La expresion genica
En todos los organismos, se requieren dos pasos para leer la información codificada en el ADN de un gen y producir la proteína que especifica. Primero, el ADN del gen se transcribe a ARN mensajero (ARNm). En segundo lugar, ese ARNm se traduce en proteína. Los genes que codifican ARN aún deben pasar por el primer paso, pero no se traducen en proteína. El proceso de producción de una molécula biológicamente funcional de ARN o proteína se denomina expresión génica, y la molécula resultante se denomina producto génico.
Codigo genetico

La secuencia de nucleótidos del ADN de un gen especifica la secuencia de aminoácidos de una proteína a través del código genético. Los conjuntos de tres nucleótidos, conocidos como codones, corresponden a un aminoácido específico. El principio de que tres bases secuenciales de código de ADN para cada aminoácido se demostró en 1961 usando mutaciones de marco de lectura en el gen rIIB del bacteriófago T4 (véase el experimento de Crick, Brenner et al.).
Además, un "codón de inicio" y tres "codones de parada" indican el comienzo y el final de la región de codificación de la proteína. Hay 64 posibles codones (cuatro posibles nucleótidos en cada una de las tres posiciones, de ahí 4 posibles codones) y solo 20 aminoácidos estándar; por lo tanto, el código es redundante y múltiples codones pueden especificar el mismo aminoácido. La correspondencia entre codones y aminoácidos es casi universal entre todos los organismos vivos conocidos.
Transcripción
La transcripción produce una molécula de ARN monocatenario conocida como ARN mensajero, cuya secuencia de nucleótidos es complementaria del ADN del que se transcribió. El ARNm actúa como un intermediario entre el gen de ADN y su producto de proteína final. El ADN del gen se usa como plantilla para generar un ARNm complementario. El ARNm coincide con la secuencia de la cadena de codificación del ADN del gen porque se sintetiza como el complemento de la cadena de la plantilla. La transcripción se realiza mediante una enzima llamada ARN polimerasa, que lee la cadena molde en la dirección 3 'a 5' y sintetiza el ARN de 5 'a 3'. Para iniciar la transcripción, la polimerasa primero reconoce y se une a una región promotora del gen. Por lo tanto, un mecanismo principal de regulación de genes es el bloqueo o secuestro de la región promotora,
En procariotas, la transcripción ocurre en el citoplasma; para transcripciones muy largas, la traducción puede comenzar en el extremo 5 'del ARN, mientras que el extremo 3' todavía se está transcribiendo. En eucariotas, la transcripción ocurre en el núcleo, donde se almacena el ADN de la célula. La molécula de ARN producida por la polimerasa se conoce como la transcripción primaria y se somete a modificaciones postranscripcionales antes de ser exportada al citoplasma para su traducción. Una de las modificaciones realizadas es el corte y empalme de intrones que son secuencias en la región transcrita que no codifican proteínas. Los mecanismos de corte y empalme alternativos pueden dar como resultado transcritos maduros del mismo gen que tienen secuencias diferentes y, de este modo, codifican proteínas diferentes. Esta es una forma importante de regulación en células eucarióticas y también ocurre en algunos procariotas.
Traducción
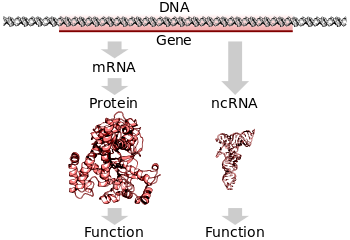
La traducción es el proceso mediante el cual se utiliza una molécula de ARNm madura como plantilla para sintetizar una nueva proteína. La traducción se lleva a cabo mediante ribosomas, grandes complejos de ARN y proteínas responsables de llevar a cabo las reacciones químicas para agregar nuevos aminoácidos a un polipéptido en crecimiento cadena por la formación de enlaces peptídicos. El código genético se lee tres nucleótidos a la vez, en unidades llamadas codones, a través de interacciones con moléculas de ARN especializadas llamadas ARN de transferencia (ARNt). Cada ARNt tiene tres bases desapareadas conocidas como el anticodón que son complementarias al codón que lee en el ARNm. El ARNt también se une covalentemente al aminoácido especificado por el codón complementario. Cuando el ARNt se une a su codón complementario en una cadena de ARNm, el ribosoma une su carga de aminoácidos a la nueva cadena polipeptídica, que se sintetiza desde el extremo amino hasta el extremo carboxilo. Durante y después de la síntesis, la mayoría de las proteínas nuevas deben plegarse a su estructura tridimensional activa antes de que puedan llevar a cabo sus funciones celulares.
Regulación
Los genes están regulados para que se expresen solo cuando se necesita el producto, ya que la expresión se basa en recursos limitados. Una célula regula su expresión génica dependiendo de su entorno externo (por ejemplo, nutrientes disponibles, temperatura y otras tensiones), su entorno interno (por ejemplo, célula ciclo de división, metabolismo, estado de infección) y su papel específico si se trata de un organismo multicelular. La expresión génica puede regularse en cualquier etapa: desde la iniciación transcripcional, hasta el procesamiento del ARN, y la modificación postraduccional de la proteína. La regulación de los genes del metabolismo de la lactosa en E. coli ( operón lac ) fue el primer mecanismo de este tipo que se describió en 1961.
Genes de ARN
Un gen codificador de proteína típico se copia primero en ARN como intermediario en la fabricación del producto de proteína final. En otros casos, las moléculas de ARN son los productos funcionales reales, como en la síntesis de ARN ribosómico y ARN de transferencia. Algunos ARN conocidos como ribozimas son capaces de funcionar enzimáticamente y los microARN tienen un papel regulador. Las secuencias de ADN a partir de las cuales se transcriben dichos ARN se conocen como genes de ARN no codificante.
Algunos virus almacenan sus genomas completos en forma de ARN y no contienen ADN en absoluto. Debido a que usan ARN para almacenar genes, sus huéspedes celulares pueden sintetizar sus proteínas tan pronto como se infectan y sin el retraso en la espera de la transcripción. Por otro lado, los retrovirus de ARN, como el VIH, requieren la transcripción inversa de su genoma del ARN al ADN antes de que sus proteínas puedan sintetizarse. La herencia epigenética mediada por ARN también se ha observado en plantas y muy raramente en animales.
Herencia

Los organismos heredan sus genes de sus padres. Los organismos asexuales simplemente heredan una copia completa del genoma de sus padres. Los organismos sexuales tienen dos copias de cada cromosoma porque heredan un conjunto completo de cada padre.
Herencia mendeliana
De acuerdo con la herencia mendeliana, las variaciones en el fenotipo de un organismo (características físicas y de comportamiento observables) se deben en parte a las variaciones en su genotipo (conjunto particular de genes). Cada gen especifica un rasgo particular con una secuencia diferente de un gen (alelos) que da lugar a diferentes fenotipos. La mayoría de los organismos eucarióticos (como las plantas de guisantes en los que trabajó Mendel) tienen dos alelos por cada rasgo, uno heredado de cada progenitor.
Los alelos en un locus pueden ser dominantes o recesivos; los alelos dominantes dan lugar a sus fenotipos correspondientes cuando se combinan con cualquier otro alelo para el mismo rasgo, mientras que los alelos recesivos dan lugar a su fenotipo correspondiente solo cuando se combinan con otra copia del mismo alelo. Si conoce los genotipos de los organismos, puede determinar qué alelos son dominantes y cuáles recesivos. Por ejemplo, si el alelo que especifica los tallos altos en plantas de arvejas es dominante sobre los tallos cortos que especifican los alelos, las plantas de guisantes que heredan un alelo alto de un padre y un alelo corto del otro progenitor también tendrán tallos altos. El trabajo de Mendel demostró que los alelos se clasifican independientemente en la producción de gametos, o células germinales, lo que garantiza la variación en la próxima generación.
Replicación de ADN y división celular
El crecimiento, desarrollo y reproducción de organismos se basa en la división celular; el proceso por el cual una sola célula se divide en dos células hijas generalmente idénticas. Esto requiere primero hacer una copia duplicada de cada gen en el genoma en un proceso llamado replicación de ADN. Las copias están hechas por enzimas especializadas conocidas como ADN polimerasas, que "leen" una cadena del ADN de doble hélice, conocida como la cadena molde, y sintetizan una nueva cadena complementaria. Debido a que la doble hélice de ADN se mantiene unida por emparejamiento de bases, la secuencia de una cadena especifica completamente la secuencia de su complemento; por lo tanto, solo una hebra necesita ser leída por la enzima para producir una copia fiel. El proceso de replicación del ADN es semiconservativo; es decir,
La tasa de replicación del ADN en células vivas se midió primero como la tasa de elongación del ADN del fago T4 en E. coli infectada con fagos y se descubrió que era impresionantemente rápida. Durante el período de aumento exponencial de ADN a 37 ° C, la tasa de elongación fue de 749 nucleótidos por segundo.
Después de completar la replicación del ADN, la célula debe separar físicamente las dos copias del genoma y dividirse en dos células distintas unidas a la membrana. En procariotas (bacterias y arqueas) esto generalmente ocurre a través de un proceso relativamente simple llamado fisión binaria, en el cual cada genoma circular se une a la membrana celular y se separa en las células hijas a medida que la membrana invagina para dividir el citoplasma en dos porciones . La fisión binaria es extremadamente rápida en comparación con las tasas de división celular en eucariotas. La división de células eucarióticas es un proceso más complejo conocido como ciclo celular; La replicación del ADN ocurre durante una fase de este ciclo conocida como fase S, mientras que el proceso de segregación de los cromosomas y la división del citoplasma ocurre durante la fase M.
Herencia molecular
La duplicación y transmisión de material genético de una generación de células a la siguiente es la base de la herencia molecular y el vínculo entre las imágenes clásicas y moleculares de los genes. Los organismos heredan las características de sus padres porque las células de la descendencia contienen copias de los genes en las células de sus padres. En organismos que se reproducen asexualmente, la descendencia será una copia genética o clon del organismo padre. En los organismos que se reproducen sexualmente, una forma especializada de división celular llamada meiosis produce células llamadas gametos o células germinales que son haploides o contienen una sola copia de cada gen. Los gametos producidos por las hembras se llaman huevos u óvulos, y los producidos por los machos se llaman espermatozoides. Dos gametos se fusionan para formar un óvulo diploide fertilizado, una única célula que tiene dos grupos de genes,
Durante el proceso de división celular meiótica, un evento llamado recombinación genética o traspaso a veces puede ocurrir, en el que una longitud de ADN en una cromátida se intercambia con una longitud de ADN en la correspondiente cromátida homóloga no hermana. Esto puede dar como resultado una recombinación de alelos enlazados de otra manera. El principio mendeliano del surtido independiente afirma que cada uno de los dos genes de un padre para cada rasgo se clasificará independientemente en gametos; el alelo que un organismo hereda para un rasgo no está relacionado con el alelo que hereda para otro rasgo. De hecho, esto solo es cierto para los genes que no residen en el mismo cromosoma o que se encuentran muy lejos el uno del otro en el mismo cromosoma. Cuanto más cerca estén dos genes en el mismo cromosoma, más estrechamente se asociarán en los gametos y con mayor frecuencia aparecerán juntos (lo que se conoce como vinculación genética).
Evolución molecular
Mutación
La replicación del ADN es, en su mayor parte, extremadamente precisa, sin embargo se producen errores (mutaciones). La tasa de error en células eucarióticas puede ser tan baja como 10 por nucleótido por replicación, mientras que para algunos virus de ARN puede ser tan alta como 10. Esto significa que cada generación, cada genoma humano acumula 1-2 nuevas mutaciones. Pequeñas mutaciones pueden ser causadas por la replicación del ADN y las secuelas del daño en el ADN e incluyen mutaciones puntuales en las que se altera una única base y mutaciones en el marco de lectura en las que se inserta o elimina una única base. Cualquiera de estas mutaciones puede cambiar el gen por missense (cambiar un codón para codificar un aminoácido diferente) o sin sentido (un codón de detención prematuro). Las mutaciones más grandes pueden ser causadas por errores en la recombinación que causan anomalías cromosómicas, incluida la duplicación, eliminación, reordenamiento o inversión de grandes secciones de un cromosoma. Además, los mecanismos de reparación del ADN pueden introducir errores mutacionales al reparar el daño físico a la molécula. La reparación, incluso con mutación, es más importante para la supervivencia que la restauración de una copia exacta, por ejemplo, al reparar roturas de doble cadena.
Cuando múltiples alelos diferentes para un gen están presentes en la población de una especie se llama polimórficos. La mayoría de los alelos diferentes son funcionalmente equivalentes, sin embargo, algunos alelos pueden dar lugar a diferentes rasgos fenotípicos. El alelo más común de un gen se llama el tipo salvaje, y los alelos raros se llaman mutantes. La variación genética en las frecuencias relativas de diferentes alelos en una población se debe tanto a la selección natural como a la deriva genética. El alelo salvaje no es necesariamente el antecesor de los alelos menos comunes, ni necesariamente es más adecuado.
La mayoría de las mutaciones dentro de los genes son neutrales, y no tienen ningún efecto sobre el fenotipo del organismo (mutaciones silenciosas). Algunas mutaciones no cambian la secuencia de aminoácidos porque codones múltiples codifican el mismo aminoácido (mutaciones sinónimas). Otras mutaciones pueden ser neutrales si conducen a cambios en la secuencia de aminoácidos, pero la proteína todavía funciona de manera similar con el nuevo aminoácido (por ejemplo, mutaciones conservadoras). Muchas mutaciones, sin embargo, son perjudiciales o incluso letales, y se eliminan de las poblaciones por selección natural. Los trastornos genéticos son el resultado de mutaciones perjudiciales y pueden deberse a una mutación espontánea en el individuo afectado, o pueden ser heredados. Finalmente, una pequeña fracción de las mutaciones es beneficiosa, mejorando la aptitud del organismo y son extremadamente importantes para la evolución,
Homología de secuencia

Los genes con un ancestro común más reciente, y por lo tanto una ascendencia evolutiva compartida, se conocen como homólogos. Estos genes aparecen por duplicación genética dentro del genoma de un organismo, donde se los conoce como genes parálogos, o son el resultado de la divergencia de los genes después de un evento de especiación, donde se los conoce como genes ortólogos y con frecuencia realizan las mismas funciones o funciones similares. en organismos relacionados. A menudo se supone que las funciones de los genes ortólogos son más similares que las de los genes parálogos, aunque la diferencia es mínima.
La relación entre los genes puede medirse comparando la alineación de secuencia de su ADN. El grado de similitud de secuencia entre genes homólogos se denomina secuencia conservada. La mayoría de los cambios en la secuencia de un gen no afectan su función, por lo que los genes acumulan mutaciones a lo largo del tiempo mediante una evolución molecular neutra. Además, cualquier selección en un gen hará que su secuencia diverja a un ritmo diferente. Los genes bajo selección estabilizadora están restringidos y cambian más lentamente mientras que los genes bajo selección direccional cambian la secuencia más rápidamente. Las diferencias de secuencia entre genes pueden usarse para análisis filogenéticos para estudiar cómo han evolucionado esos genes y cómo se relacionan los organismos de los que provienen.
Orígenes de nuevos genes

La fuente más común de nuevos genes en linajes eucarióticos es la duplicación de genes, que crea la variación del número de copias de un gen existente en el genoma. Los genes resultantes (parálogos) pueden divergir en secuencia y en función. Conjuntos de genes formados de esta manera componen una familia de genes. Las duplicaciones y pérdidas genéticas dentro de una familia son comunes y representan una fuente importante de biodiversidad evolutiva. A veces, la duplicación de genes puede dar como resultado una copia no funcional de un gen, o una copia funcional puede estar sujeta a mutaciones que dan como resultado la pérdida de la función; tales genes no funcionales se llaman pseudogenes.
La transferencia horizontal de genes se refiere a la transferencia de material genético a través de un mecanismo distinto de la reproducción. Este mecanismo es una fuente común de nuevos genes en procariotas, a veces se cree que contribuyen más a la variación genética que a la duplicación de genes. Es un medio común de propagar la resistencia a los antibióticos, la virulencia y las funciones metabólicas adaptativas. Aunque la transferencia horizontal de genes es rara en eucariotas, se han identificado ejemplos probables de protistas y algagenomas que contienen genes de origen bacteriano.
Genoma
El genoma es el material genético total de un organismo e incluye tanto los genes como las secuencias no codificantes.
Numero de genes

El tamaño del genoma y el número de genes que codifica varían ampliamente entre los organismos. Los genomas más pequeños se encuentran en virus y viroides (que actúan como un solo gen de ARN no codificante). Por el contrario, las plantas pueden tener genomas extremadamente grandes, con arroz que contiene> 46,000 genes que codifican proteínas. Se estima que el número total de genes que codifican proteínas (el proteoma de la Tierra) es de 5 millones de secuencias.
Aunque el número de pares de bases de ADN en el genoma humano se conoce desde la década de 1960, el número estimado de genes ha cambiado con el tiempo a medida que se han ido perfeccionando las definiciones de los genes y los métodos para detectarlos. Las predicciones teóricas iniciales del número de genes humanos fueron tan altas como 2,000,000. Las primeras medidas experimentales indicaban que había de 50,000 a 100,000 transcripciones genes (etiquetas de secuencias expresadas). Posteriormente, la secuenciación en el Proyecto Genoma Humano indicó que muchas de estas transcripciones eran variantes de los mismos genes, y el número total de genes que codifican proteínas se modificó a ~ 20,000 con 13 genes codificados en el genoma mitocondrial. Con el proyecto de anotación GENCODE, ese cálculo ha seguido cayendo a 19,000. Del genoma humano, solo el 1-2% consiste en genes que codifican proteínas, y el resto es ADN "no codificante", como intrones, retrotransposones y ARN no codificantes. Cada organismo multicelular tiene todos sus genes en cada célula de su cuerpo, pero no todas las funciones de genes en cada célula.
Genes esenciales
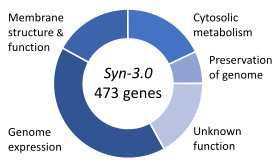
Los genes esenciales son el conjunto de genes considerados críticos para la supervivencia de un organismo. Esta definición supone la disponibilidad abundante de todos los nutrientes relevantes y la ausencia de estrés ambiental. Solo una pequeña porción de los genes de un organismo es esencial. En las bacterias, se estima que 250-400 genes son esenciales para Escherichia coli y Bacillus subtilis , que es menos del 10% de sus genes. La mitad de estos genes son ortólogos en ambos organismos y están muy involucrados en la síntesis de proteínas. En la levadura en ciernes Saccharomyces cerevisiaeel número de genes esenciales es ligeramente más alto, en 1000 genes (~ 20% de sus genes). Aunque el número es más difícil de medir en eucariotas superiores, se estima que los ratones y los humanos tienen alrededor de 2000 genes esenciales (~ 10% de sus genes). El organismo sintético, Syn 3 , tiene un genoma mínimo de 473 genes esenciales y genes casi esenciales (necesarios para un crecimiento rápido), aunque 149 tienen una función desconocida.
Los genes esenciales incluyen genes de limpieza (esenciales para las funciones celulares básicas), así como genes que se expresan en diferentes momentos en el desarrollo de los organismos o el ciclo de vida. Los genes de limpieza se utilizan como controles experimentales al analizar la expresión génica, ya que se expresan constitutivamente a un nivel relativamente constante.
Nomenclatura genética y genómica
La nomenclatura de genes ha sido establecida por el HUGO Gene Nomenclature Committee (HGNC) para cada gen humano conocido en la forma de un nombre genético aprobado y un símbolo (abreviatura abreviada), al que se puede acceder a través de una base de datos mantenida por HGNC. Los símbolos se eligen para ser únicos, y cada gen tiene solo un símbolo (aunque los símbolos aprobados a veces cambian). Los símbolos se mantienen preferiblemente consistentes con otros miembros de una familia de genes y con homólogos en otras especies, particularmente el ratón debido a su papel como organismo modelo común.
Ingeniería genética
La ingeniería genética es la modificación del genoma de un organismo a través de la biotecnología. Desde la década de 1970, se han desarrollado una variedad de técnicas para agregar, eliminar y editar genes específicamente en un organismo. Las técnicas de ingeniería del genoma recientemente desarrolladas usan enzimas nucleasa diseñadas para crear una reparación específica del ADN en un cromosoma para interrumpir o editar un gen cuando se repara la ruptura. El término relacionado biología sintética se usa a veces para referirse a la ingeniería genética extensa de un organismo.
La ingeniería genética es ahora una herramienta de investigación de rutina con organismos modelo. Por ejemplo, los genes se agregan fácilmente a las bacterias y los linajes de ratones knockout con la función de un gen específico alterado se utilizan para investigar la función de ese gen. Muchos organismos han sido genéticamente modificados para aplicaciones en agricultura, biotecnología industrial y medicina.
Para los organismos multicelulares, generalmente el embrión está diseñado, que crece en el organismo genéticamente modificado adulto. Sin embargo, los genomas de las células en un organismo adulto se pueden editar utilizando técnicas de terapia génica para tratar enfermedades genéticas.