Genética
Definición
La genética es el estudio de los genes, la variación genética y la herencia en los organismos vivos. En general, se considera un campo de la biología, pero se cruza con frecuencia con muchas otras ciencias de la vida y está fuertemente relacionado con el estudio de los sistemas de información.El descubridor de la genética es Gregor Mendel, un científico de finales del siglo XIX y fraile agustino. Mendel estudió "herencia de rasgos", patrones en la forma en que los rasgos se transmiten de padres a hijos. Observó que los organismos (plantas de guisantes) heredan rasgos a través de "unidades de herencia" discretas. Este término, que aún se usa hoy en día, es una definición algo ambigua de lo que se conoce como un gen.
La herencia de rasgos y los mecanismos de herencia molecular de los genes siguen siendo los principios primarios de la genética en el siglo XXI, pero la genética moderna se ha expandido más allá de la herencia para estudiar la función y el comportamiento de los genes. La estructura y la función del gen, la variación y la distribución se estudian en el contexto de la célula, el organismo (por ejemplo, el dominio) y dentro del contexto de una población. La genética ha dado lugar a una serie de subcampos, incluida la epigenética y la genética de poblaciones. Los organismos estudiados dentro del amplio campo abarcan los dominios de la vida (arqueas, bacterias y eukarya).
Los procesos genéticos funcionan en combinación con el entorno y las experiencias de un organismo para influir en el desarrollo y el comportamiento, a menudo denominado naturaleza versus crianza. El entorno intracelular o extracelular de una célula u organismo puede activar o desactivar la transcripción génica. Un ejemplo clásico es dos semillas de maíz genéticamente idéntico, uno colocado en un clima templado y otro en un clima árido. Si bien la altura promedio de los dos tallos de maíz puede determinarse genéticamente para ser igual, la del clima árido solo crece a la mitad de la que tiene el clima templado debido a la falta de agua y nutrientes en su entorno.
Etimología
La palabra genética proviene del griego antiguo γενετικός genetikos que significa "genitivo" / "generativo", que a su vez deriva de γένεσις génesis que significa "origen".
Historia
La observación de que los seres vivos heredan rasgos de sus padres se ha utilizado desde tiempos prehistóricos para mejorar las plantas y los animales de cultivo mediante la cría selectiva. La ciencia moderna de la genética, que busca comprender este proceso, comenzó con el trabajo del fraile agustino Gregor Mendel a mediados del siglo XIX.
Antes de Mendel, Imre Festetics, un noble húngaro, que vivió en Kőszeg antes que Mendel, fue el primero en usar la palabra "genética". Describió varias reglas de la herencia genética en su obra La ley genética de la naturaleza (Die genetische Gesätze der Natur, 1819). Su segunda ley es la misma que Mendel publicó. En su tercera ley, desarrolló los principios básicos de la mutación (puede considerarse un precursor de Hugo de Vries).

Otras teorías de herencia precedieron el trabajo de Mendel. Una teoría popular durante el siglo XIX, e implícita en el libro de Charles Darwin de 1859 Sobre el origen de las especies, estaba mezclando herencia: la idea de que los individuos heredan una mezcla suave de rasgos de sus padres. El trabajo de Mendel proporcionó ejemplos donde los rasgos definitivamente no se mezclaron después de la hibridación, lo que demuestra que los rasgos se producen mediante combinaciones de genes distintos en lugar de una mezcla continua. La combinación de rasgos en la progenie ahora se explica por la acción de múltiples genes con efectos cuantitativos. Otra teoría que tuvo cierto apoyo en ese momento fue la herencia de las características adquiridas: la creencia de que los individuos heredan rasgos fortalecidos por sus padres. Se sabe que esta teoría (comúnmente asociada con Jean-Baptiste Lamarck) es errónea: las experiencias de los individuos no afectan los genes que transmiten a sus hijos, aunque la evidencia en el campo de la epigenética ha revivido algunos aspectos de la teoría de Lamarck.
Genética mendeliana y clásica

La genética moderna comenzó con los estudios de Mendel sobre la naturaleza de la herencia en las plantas. En su artículo " Versuche über Pflanzenhybriden " ("Experimentos sobre hibridación de plantas"), presentado en 1865 a la Naturforschender Verein (Sociedad para la Investigación de la Naturaleza) en Brünn, Mendel trazó los patrones de herencia de ciertos rasgos en plantas de guisantes y los describió matemáticamente. Aunque este patrón de herencia solo se podía observar para algunos rasgos, el trabajo de Mendel sugería que la herencia era particulada, no adquirida, y que los patrones de herencia de muchos rasgos podían explicarse mediante reglas y proporciones simples.
La importancia del trabajo de Mendel no obtuvo una amplia comprensión hasta 1900, después de su muerte, cuando Hugo de Vries y otros científicos redescubrieron su investigación. William Bateson, un defensor del trabajo de Mendel, acuñó la palabra genética en 1905 (el adjetivo genético , derivado de la palabra griega genesis -γένεσις, "origen", es anterior al sustantivo y se utilizó por primera vez en un sentido biológico en 1860). Bateson actuó como mentor y fue ayudado significativamente por el trabajo de mujeres científicas del Newnham College en Cambridge, específicamente el trabajo de Becky Saunders, Nora Darwin Barlow y Muriel Wheldale Onslow. Bateson popularizó el uso de la palabra genética para describir el estudio de la herencia en su discurso inaugural de la Tercera Conferencia Internacional sobre Hibridación de Plantas en Londres en 1906.
Después del redescubrimiento del trabajo de Mendel, los científicos trataron de determinar qué moléculas de la célula eran responsables de la herencia. En 1911, Thomas Hunt Morgan argumentó que los genes están en los cromosomas, en base a las observaciones de una mutación del ojo blanco ligada al sexo en las moscas de la fruta. En 1913, su alumno Alfred Sturtevant utilizó el fenómeno de la vinculación genética para mostrar que los genes están dispuestos linealmente en el cromosoma.
Genética molecular
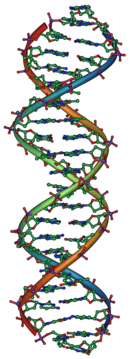
Aunque se sabía que los genes existían en los cromosomas, los cromosomas están compuestos de proteína y ADN, y los científicos no sabían cuál de los dos es responsable de la herencia. En 1928, Frederick Griffith descubrió el fenómeno de la transformación (ver el experimento de Griffith): las bacterias muertas podían transferir material genético para "transformar" otras bacterias que aún vivían. Dieciséis años más tarde, en 1944, el experimento Avery-MacLeod-McCarty identificó al ADN como la molécula responsable de la transformación. El papel del núcleo como depósito de información genética en eucariotas había sido establecido por Hämmerling en 1943 en su trabajo sobre el alga unicelular Acetabularia. El experimento de Hershey-Chase en 1952 confirmó que el ADN (en lugar de la proteína) es el material genético de los virus que infectan a las bacterias, lo que proporciona más evidencia de que el ADN es la molécula responsable de la herencia.
James Watson y Francis Crick determinaron la estructura del ADN en 1953, utilizando el trabajo de cristalografía de rayos X de Rosalind Franklin y Maurice Wilkins que indicaba que el ADN tiene una estructura helicoidal (es decir, tiene la forma de un sacacorchos). Su modelo de doble hélice tenía dos cadenas de ADN con los nucleótidos apuntando hacia adentro, cada uno haciendo coincidir un nucleótido complementario en la otra cadena para formar lo que parecen peldaños en una escalera retorcida. Esta estructura mostró que existe información genética en la secuencia de nucleótidos en cada cadena de ADN. La estructura también sugirió un método simple para la replicación: si los filamentos están separados, se pueden reconstruir nuevos hilos asociados para cada uno en función de la secuencia de la cadena anterior. Esta propiedad es lo que le da al ADN su naturaleza semi-conservativa en la que una cadena de ADN nuevo proviene de una cadena matriz original.
Aunque la estructura del ADN mostró cómo funciona la herencia, aún no se sabía cómo el ADN influye en el comportamiento de las células. En los años siguientes, los científicos trataron de comprender cómo el ADN controla el proceso de producción de proteínas. Se descubrió que la célula usa ADN como plantilla para crear ARN mensajero correspondiente, moléculas con nucleótidos muy similares al ADN. La secuencia de nucleótidos de un ARN mensajero se usa para crear una secuencia de aminoácidos en la proteína; esta traducción entre secuencias de nucleótidos y secuencias de aminoácidos se conoce como el código genético.
Con la nueva comprensión molecular de la herencia surgió una explosión de investigación. Una teoría notable surgió de Tomoko Ohta en 1973 con su enmienda a la teoría neutral de la evolución molecular mediante la publicación de la teoría casi neutral de la evolución molecular. En esta teoría, Ohta hizo hincapié en la importancia de la selección natural y el medio ambiente a la velocidad a la que se produce la evolución genética. Un desarrollo importante fue la secuenciación de ADN de terminación de cadena en 1977 por Frederick Sanger. Esta tecnología permite a los científicos leer la secuencia de nucleótidos de una molécula de ADN. En 1983, Kary Banks Mullis desarrolló la reacción en cadena de la polimerasa, proporcionando una forma rápida de aislar y amplificar una sección específica del ADN de una mezcla. Los esfuerzos del Proyecto Genoma Humano, Departamento de Energía, NIH,
Características de la herencia
Herencia discreta y las leyes de Mendel
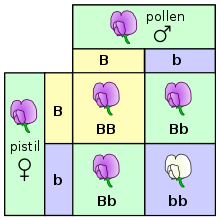
En su nivel más fundamental, la herencia en los organismos ocurre al pasar unidades heredables discretas, llamadas genes, de padres a hijos. Esta propiedad fue observada por primera vez por Gregor Mendel, quien estudió la segregación de los rasgos hereditarios en las plantas de guisantes. En sus experimentos estudiando el rasgo del color de la flor, Mendel observó que las flores de cada planta de guisante eran de color púrpura o blanco, pero nunca intermedias entre los dos colores. Estas diferentes versiones discretas del mismo gen se llaman alelos.
En el caso del guisante, que es una especie diploide, cada planta individual tiene dos copias de cada gen, una copia heredada de cada padre. Muchas especies, incluidos los humanos, tienen este patrón de herencia. Los organismos diploides con dos copias del mismo alelo de un gen dado se llaman homocigotos en ese locus del gen, mientras que los organismos con dos alelos diferentes de un gen dado se denominan heterocigotos.
El conjunto de alelos para un organismo dado se llama su genotipo, mientras que los rasgos observables del organismo se llaman su fenotipo. Cuando los organismos son heterocigotos en un gen, a menudo un alelo se denomina dominante ya que sus cualidades dominan el fenotipo del organismo, mientras que el otro alelo se denomina recesivo a medida que sus cualidades retroceden y no se observan. Algunos alelos no tienen dominancia completa y en su lugar tienen una dominancia incompleta al expresar un fenotipo intermedio, o codominancia al expresar ambos alelos a la vez.
Cuando un par de organismos se reproducen sexualmente, sus descendientes heredan aleatoriamente uno de los dos alelos de cada padre. Estas observaciones de la herencia discreta y la segregación de los alelos se conocen colectivamente como la primera ley de Mendel o la Ley de la segregación.
Notación y diagramas

Los genetistas usan diagramas y símbolos para describir la herencia. Un gen está representado por una o algunas letras. Con frecuencia, se usa un símbolo "+" para marcar el alelo habitual, no mutante, de un gen.
En experimentos de fertilización y reproducción (y especialmente cuando se discuten las leyes de Mendel) se hace referencia a los padres como la generación "P" y la descendencia como la generación "F1" (primera filial). Cuando la descendencia F1 se aparean entre sí, la descendencia recibe el nombre de generación "F2" (segunda filial). Uno de los diagramas comunes usados para predecir el resultado del cruce es el cuadro de Punnett.
Cuando se estudian las enfermedades genéticas humanas, los genetistas a menudo usan gráficas de pedigrí para representar la herencia de los rasgos. Estas tablas mapean la herencia de un rasgo en un árbol genealógico.
Interacciones genéticas múltiples
Los organismos tienen miles de genes y, en los organismos que se reproducen sexualmente, estos genes generalmente se unen de forma independiente. Esto significa que la herencia de un alelo de color amarillo o verde no está relacionada con la herencia de alelos para flores blancas o moradas. Este fenómeno, conocido como la "segunda ley de Mendel" o la "ley del surtido independiente", significa que los alelos de diferentes genes se mezclan entre los padres para formar descendencia con muchas combinaciones diferentes. (Algunos genes no se clasifican de forma independiente, lo que demuestra la vinculación genética, un tema que se analiza más adelante en este artículo).
A menudo, diferentes genes pueden interactuar de una manera que influye en el mismo rasgo. En la María de ojos azules ( Omphalodes verna ), por ejemplo, existe un gen con alelos que determinan el color de las flores: azul o magenta. Otro gen, sin embargo, controla si las flores tienen color o si son blancas. Cuando una planta tiene dos copias de este alelo blanco, sus flores son blancas, independientemente de si el primer gen tiene alelos azul o magenta. Esta interacción entre los genes se llama epistasis, con el segundo gen epistatic a la primera.
Muchos rasgos no son características discretas (por ejemplo, flores púrpuras o blancas) sino que son características continuas (por ejemplo, altura humana y color de piel). Estos rasgos complejos son productos de muchos genes. La influencia de estos genes está mediada, en diversos grados, por el entorno que ha experimentado un organismo. El grado en que los genes de un organismo contribuyen a un rasgo complejo se llama heredabilidad. La medición de la heredabilidad de un rasgo es relativa: en un entorno más variable, el ambiente tiene una mayor influencia en la variación total del rasgo. Por ejemplo, la altura humana es un rasgo con causas complejas. Tiene una heredabilidad del 89% en los Estados Unidos. En Nigeria, sin embargo, donde las personas experimentan un acceso más variable a una buena nutrición y cuidado de la salud, la altura tiene una heredabilidad de solo el 62%.
Bases moleculares para la herencia
ADN y cromosomas

La base molecular de los genes es el ácido desoxirribonucleico (ADN). El ADN se compone de una cadena de nucleótidos, de los cuales hay cuatro tipos: adenina (A), citosina (C), guanina (G) y timina (T). La información genética existe en la secuencia de estos nucleótidos, y los genes existen como tramos de secuencia a lo largo de la cadena de ADN. Los virus son la única excepción a esta regla: a veces los virus usan una molécula de ARN muy similar en lugar de ADN como material genético. Los virus no pueden reproducirse sin un huésped y no se ven afectados por muchos procesos genéticos, por lo que tienden a no ser considerados organismos vivos.
El ADN normalmente existe como una molécula bicatenaria, enrollada en forma de doble hélice. Cada nucleótido en el ADN se empareja preferentemente con su nucleótido compañero en la cadena opuesta: A se empareja con T y C se empareja con G. Por lo tanto, en su forma bicatenaria, cada cadena contiene efectivamente toda la información necesaria, redundante con su cadena compañera. Esta estructura del ADN es la base física de la herencia: la replicación del ADN duplica la información genética al dividir los filamentos y utilizar cada cadena como plantilla para la síntesis de una nueva cadena asociada.
Los genes están dispuestos linealmente a lo largo de largas cadenas de secuencias de pares de bases de ADN. En las bacterias, cada célula generalmente contiene un único genophore circular, mientras que los organismos eucarióticos (como las plantas y los animales) tienen su ADN dispuesto en múltiples cromosomas lineales. Estas cadenas de ADN a menudo son extremadamente largas; el cromosoma humano más grande, por ejemplo, tiene aproximadamente 247 millones de pares de bases de longitud. El ADN de un cromosoma se asocia con proteínas estructurales que organizan, compactan y controlan el acceso al ADN, formando un material llamado cromatina; en eucariotas, la cromatina generalmente está compuesta de nucleosomas, segmentos de ADN enrollados alrededor de núcleos de proteínas de histonas. El conjunto completo de material hereditario en un organismo (generalmente las secuencias de ADN combinadas de todos los cromosomas) se denomina genoma.
Mientras que los organismos haploides tienen una sola copia de cada cromosoma, la mayoría de los animales y muchas plantas son diploides, que contienen dos de cada cromosoma y, por lo tanto, dos copias de cada gen. Los dos alelos para un gen se encuentran en loci idénticos de los dos cromosomas homólogos, cada alelo heredado de un padre diferente.

Muchas especies tienen los llamados cromosomas sexuales que determinan el género de cada organismo. En humanos y en muchos otros animales, el cromosoma Y contiene el gen que desencadena el desarrollo de las características específicamente masculinas. En la evolución, este cromosoma ha perdido la mayor parte de su contenido y también la mayoría de sus genes, mientras que el cromosoma X es similar a los otros cromosomas y contiene muchos genes. Los cromosomas X e Y forman un par muy heterogéneo.
Reproducción
Cuando las células se dividen, se copia su genoma completo y cada célula hija hereda una copia. Este proceso, llamado mitosis, es la forma más simple de reproducción y es la base de la reproducción asexual. La reproducción asexual también puede ocurrir en organismos multicelulares, produciendo descendientes que heredan su genoma de un padre soltero. Los descendientes que son genéticamente idénticos a sus padres se llaman clones.
Los organismos eucariotas a menudo usan la reproducción sexual para generar descendencia que contiene una mezcla de material genético heredado de dos padres diferentes. El proceso de reproducción sexual alterna entre formas que contienen copias únicas del genoma (haploide) y copias dobles (diploides). Las células haploides fusionan y combinan material genético para crear una célula diploide con cromosomas emparejados. Los organismos diploides forman haploides al dividirse, sin replicar su ADN, para crear células hijas que heredan aleatoriamente uno de cada par de cromosomas. La mayoría de los animales y muchas plantas son diploides durante la mayor parte de su vida, con la forma haploide reducida a gametos de una sola célula, como la esperma o los huevos.
Aunque no usan el método de reproducción sexual haploide / diploide, las bacterias tienen muchos métodos para adquirir nueva información genética. Algunas bacterias pueden experimentar conjugación, transfiriendo una pequeña porción circular de ADN a otra bacteria. Las bacterias también pueden tomar fragmentos de ADN en bruto que se encuentran en el ambiente e integrarlos en sus genomas, un fenómeno conocido como transformación. Estos procesos resultan en la transferencia horizontal de genes, transmitiendo fragmentos de información genética entre organismos que de otro modo no estarían relacionados.
Recombinación y vinculación genética

La naturaleza diploide de los cromosomas permite que los genes en diferentes cromosomas se clasifiquen independientemente o se separen de su pareja homóloga durante la reproducción sexual en la que se forman los gametos haploides. De esta forma, pueden aparecer nuevas combinaciones de genes en la descendencia de un par de apareamiento. Los genes en el mismo cromosoma teóricamente nunca se recombinarían. Sin embargo, lo hacen, a través del proceso celular de cruce cromosómico. Durante el cruce, los cromosomas intercambian tramos de ADN, mezclando los alelos genéticos entre los cromosomas. Este proceso de cruce cromosómico generalmente ocurre durante la meiosis, una serie de divisiones celulares que crean células haploides.
La primera demostración citológica de sobrecruzamiento fue realizada por Harriet Creighton y Barbara McClintock en 1931. Su investigación y experimentos en maíz proporcionaron evidencia citológica de la teoría genética que vinculaba genes en cromosomas pareados, de hecho, intercambiaban lugares de un homólogo al otro.
La probabilidad de cruce cromosómico que ocurre entre dos puntos dados en el cromosoma se relaciona con la distancia entre los puntos. Para una distancia arbitrariamente larga, la probabilidad de cruce es lo suficientemente alta como para que la herencia de los genes no esté correlacionada. Para los genes que están más juntos, sin embargo, la menor probabilidad de cruce significa que los genes demuestran la vinculación genética; los alelos para los dos genes tienden a heredarse juntos. Las cantidades de enlace entre una serie de genes se pueden combinar para formar un mapa de enlace lineal que describe aproximadamente la disposición de los genes a lo largo del cromosoma.
La expresion genica
Codigo genetico

Los genes generalmente expresan su efecto funcional a través de la producción de proteínas, que son moléculas complejas responsables de la mayoría de las funciones en la célula. Las proteínas están compuestas por una o más cadenas de polipéptidos, cada una de las cuales está compuesta por una secuencia de aminoácidos, y la secuencia de ADN de un gen (a través de un ARN intermedio) se usa para producir una secuencia de aminoácidos específica. Este proceso comienza con la producción de una molécula de ARN con una secuencia que coincide con la secuencia de ADN del gen, un proceso llamado transcripción.
Esta molécula de ARN mensajero se usa para producir una secuencia de aminoácidos correspondiente a través de un proceso llamado traducción. Cada grupo de tres nucleótidos en la secuencia, llamado codón, corresponde a uno de los veinte posibles aminoácidos en una proteína o una instrucción para finalizar la secuencia de aminoácidos; esta correspondencia se llama código genético. El flujo de información es unidireccional: la información se transfiere de las secuencias de nucleótidos a la secuencia de aminoácidos de las proteínas, pero nunca se transfiere de la proteína a la secuencia del ADN, un fenómeno que Francis Crick llamó el dogma central de la biología molecular.
La secuencia específica de aminoácidos da como resultado una estructura tridimensional única para esa proteína, y las estructuras tridimensionales de las proteínas se relacionan con sus funciones. Algunos son moléculas estructurales simples, como las fibras formadas por el colágeno proteico. Las proteínas se pueden unir a otras proteínas y moléculas simples, a veces actuando como enzimas al facilitar las reacciones químicas dentro de las moléculas unidas (sin cambiar la estructura de la proteína en sí). La estructura de la proteína es dinámica; la proteína hemoglobina se dobla en formas ligeramente diferentes ya que facilita la captura, el transporte y la liberación de moléculas de oxígeno dentro de la sangre de mamíferos.
Una única diferencia de nucleótidos dentro del ADN puede causar un cambio en la secuencia de aminoácidos de una proteína. Debido a que las estructuras de proteínas son el resultado de sus secuencias de aminoácidos, algunos cambios pueden cambiar drásticamente las propiedades de una proteína al desestabilizar la estructura o al cambiar la superficie de la proteína de una manera que cambia su interacción con otras proteínas y moléculas. Por ejemplo, la anemia de células falciformes es una enfermedad genética humana que resulta de una única diferencia de bases dentro de la región de codificación para la sección de la globina β de la hemoglobina, lo que causa un cambio de aminoácido único que cambia las propiedades físicas de la hemoglobina. Las versiones de células falciformes de la hemoglobina se adhieren a sí mismas, apiladas para formar fibras que distorsionan la forma de los glóbulos rojos que transportan la proteína. Estas células en forma de hoz ya no fluyen suavemente a través de los vasos sanguíneos,
Algunas secuencias de ADN se transcriben en ARN, pero no se traducen en productos proteicos, tales moléculas de ARN se llaman ARN no codificante. En algunos casos, estos productos se pliegan en estructuras que están involucradas en funciones celulares críticas (por ejemplo, ARN ribosómico y ARN de transferencia). El ARN también puede tener efectos reguladores a través de interacciones de hibridación con otras moléculas de ARN (por ejemplo, ARN micro).
Naturaleza y educación

Aunque los genes contienen toda la información que un organismo utiliza para funcionar, el medio ambiente juega un papel importante en la determinación de los fenotipos finales que muestra un organismo. La frase "naturaleza y nutrición" se refiere a esta relación complementaria. El fenotipo de un organismo depende de la interacción de los genes y el medio ambiente. Un ejemplo interesante es la coloración del pelaje del gato siamés. En este caso, la temperatura corporal del gato juega el papel del ambiente. Los genes del gato codifican el cabello oscuro, por lo tanto, las células productoras de pelo en el gato producen proteínas celulares que producen cabello oscuro. Pero estas proteínas productoras de pelo oscuro son sensibles a la temperatura (es decir, tienen una mutación que causa sensibilidad a la temperatura) y se desnaturalizan en ambientes con temperaturas más altas, no producir pigmento de pelo oscuro en áreas donde el gato tiene una temperatura corporal más alta. En un ambiente de baja temperatura, sin embargo, la estructura de la proteína es estable y produce pigmento de cabello oscuro normalmente. La proteína sigue siendo funcional en áreas de la piel que son más frías, como las piernas, las orejas, la cola y la cara, por lo que el gato tiene el pelo oscuro en sus extremidades.
El medio ambiente juega un papel importante en los efectos de la fenilcetonuria de la enfermedad genética humana. La mutación que causa la fenilcetonuria interrumpe la capacidad del cuerpo para descomponer el aminoácido fenilalanina, causando una acumulación tóxica de una molécula intermedia que, a su vez, causa síntomas graves de discapacidad intelectual progresiva y convulsiones. Sin embargo, si alguien con la mutación de fenilcetonuria sigue una dieta estricta que evita este aminoácido, se mantienen normales y saludables.
Un método común para determinar cómo los genes y el medio ambiente ("naturaleza y nutrición") contribuyen a un fenotipo implica el estudio de gemelos idénticos y fraternales, u otros hermanos de nacimientos múltiples. Debido a que hermanos idénticos provienen del mismo cigoto, son genéticamente iguales. Los gemelos fraternos son genéticamente diferentes el uno del otro como hermanos normales. Al comparar la frecuencia con que ocurre un determinado trastorno en un par de gemelos idénticos con la frecuencia con que ocurre en un par de gemelos fraternos, los científicos pueden determinar si ese trastorno es causado por factores ambientales genéticos o posnatales, ya sea que tenga "naturaleza" o "nurture" "causas" Un ejemplo famoso involucró el estudio de los cuatrillizos de Genain, que eran cuatrillizos idénticos a todos diagnosticados con esquizofrenia.
Regulación genética
El genoma de un organismo dado contiene miles de genes, pero no todos estos genes necesitan estar activos en un momento dado. Un gen se expresa cuando se transcribe en ARNm y existen muchos métodos celulares para controlar la expresión de genes de modo que las proteínas se producen solo cuando las necesita la célula. Los factores de transcripción son proteínas reguladoras que se unen al ADN, ya sea promoviendo o inhibiendo la transcripción de un gen. Dentro del genoma de Escherichia coli bacterias, por ejemplo, existe una serie de genes necesarios para la síntesis del aminoácido triptófano. Sin embargo, cuando el triptófano ya está disponible para la célula, estos genes para la síntesis de triptófano ya no son necesarios. La presencia de triptófano afecta directamente a la actividad de los genes: las moléculas de triptófano se unen al represor de triptófano (un factor de transcripción), cambiando la estructura del represor de manera que el represor se une a los genes. El represor de triptófano bloquea la transcripción y la expresión de los genes, creando así una regulación de retroalimentación negativa del proceso de síntesis de triptófano.
Las diferencias en la expresión génica son especialmente claras dentro de los organismos multicelulares, donde todas las células contienen el mismo genoma pero tienen estructuras y comportamientos muy diferentes debido a la expresión de diferentes conjuntos de genes. Todas las células de un organismo multicelular se derivan de una única célula, que se diferencian en tipos de células variantes en respuesta a señales externas e intercelulares y establecen gradualmente diferentes patrones de expresión génica para crear diferentes comportamientos. Como ningún gen es responsable del desarrollo de estructuras dentro de organismos multicelulares, estos patrones surgen de las interacciones complejas entre muchas células.
Dentro de los eucariotas, existen características estructurales de la cromatina que influyen en la transcripción de genes, a menudo en forma de modificaciones en el ADN y la cromatina que son heredadas de manera estable por las células hijas. Estas características se llaman "epigenéticas" porque existen "en la parte superior" de la secuencia de ADN y retienen la herencia de una generación de células a la siguiente. Debido a las características epigenéticas, diferentes tipos de células cultivadas en el mismo medio pueden retener propiedades muy diferentes. Aunque las características epigenéticas son generalmente dinámicas en el curso del desarrollo, algunas, como el fenómeno de la paramutación, tienen herencia multigeneracional y existen como raras excepciones a la regla general del ADN como base de la herencia.
Cambio genético
Mutaciones

Durante el proceso de replicación del ADN, ocasionalmente ocurren errores en la polimerización de la segunda cadena. Estos errores, llamados mutaciones, pueden afectar el fenotipo de un organismo, especialmente si se producen dentro de la secuencia codificante de la proteína de un gen. Las tasas de error suelen ser muy bajas: 1 error en cada 10-100 millones de bases, debido a la capacidad de "revisión" de las ADN polimerasas. Los procesos que aumentan la tasa de cambios en el ADN se denominan mutagénicos: los productos químicos mutagénicos promueven errores en la replicación del ADN, a menudo interfiriendo con la estructura del apareamiento de bases, mientras que la radiación UV induce mutaciones al dañar la estructura del ADN. El daño químico al ADN también ocurre naturalmente y las células usan mecanismos de reparación del ADN para reparar los desajustes y rupturas. La reparación, sin embargo, no siempre restaura la secuencia original.
En los organismos que usan el cruce cromosómico para intercambiar ADN y recombinar genes, los errores en la alineación durante la meiosis también pueden causar mutaciones. Los errores en el cruce son especialmente probables cuando secuencias similares causan que los cromosomas asociados adopten una alineación errónea; esto hace que algunas regiones en genomas sean más propensas a mutar de esta manera. Estos errores crean grandes cambios estructurales en la secuencia de ADN (duplicaciones, inversiones, eliminaciones de regiones enteras) o el intercambio accidental de partes enteras de secuencias entre cromosomas diferentes (translocación cromosómica).

Selección natural y evolución
Las mutaciones alteran el genotipo de un organismo y ocasionalmente esto hace que aparezcan diferentes fenotipos. La mayoría de las mutaciones tienen poco efecto sobre el fenotipo, la salud o la capacidad reproductiva de un organismo. Las mutaciones que tienen un efecto suelen ser perjudiciales, pero ocasionalmente algunas pueden ser beneficiosas. Los estudios en la mosca Drosophila melanogaster sugieren que si una mutación cambia una proteína producida por un gen, aproximadamente el 70 por ciento de estas mutaciones serán perjudiciales y el resto será neutral o poco beneficioso.

La genética de poblaciones estudia la distribución de las diferencias genéticas dentro de las poblaciones y cómo estas distribuciones cambian con el tiempo. Los cambios en la frecuencia de un alelo en una población están influenciados principalmente por la selección natural, donde un alelo determinado proporciona una ventaja selectiva o reproductiva para el organismo, así como otros factores como mutación, deriva genética, autostop genético, selección artificial y migración.
Durante muchas generaciones, los genomas de los organismos pueden cambiar significativamente, lo que resulta en la evolución. En el proceso llamado adaptación, la selección de mutaciones benéficas puede hacer que una especie evolucione hacia formas más capaces de sobrevivir en su entorno. Las nuevas especies se forman mediante el proceso de especiación, a menudo causado por separaciones geográficas que impiden que las poblaciones intercambien genes entre sí.
Al comparar la homología entre los genomas de diferentes especies, es posible calcular la distancia evolutiva entre ellos y cuándo pueden haber divergido. Las comparaciones genéticas generalmente se consideran un método más preciso para caracterizar la relación entre especies que la comparación de características fenotípicas. Las distancias evolutivas entre especies se pueden usar para formar árboles evolutivos; estos árboles representan el descenso y la divergencia común de especies a lo largo del tiempo, aunque no muestran la transferencia de material genético entre especies no relacionadas (conocida como transferencia horizontal de genes y más común en bacterias).
Organismos modelo

Aunque los genetistas originalmente estudiaron la herencia en una amplia gama de organismos, los investigadores comenzaron a especializarse en el estudio de la genética de un subconjunto particular de organismos. El hecho de que ya existe una investigación significativa para un organismo dado alentaría a los nuevos investigadores a elegirla para su posterior estudio, y así, finalmente, algunos organismos modelo se convirtieron en la base de la mayoría de la investigación genética. Los temas de investigación comunes en genética de organismos modelo incluyen el estudio de la regulación de genes y la participación de genes en el desarrollo y el cáncer.
Los organismos fueron elegidos, en parte, por conveniencia: tiempos de generación cortos y la manipulación genética fácil hizo que algunos organismos fueran herramientas populares de investigación genética. Los organismos modelo ampliamente utilizados incluyen la bacteria intestinal Escherichia coli , la planta Arabidopsis thaliana , la levadura de panadería ( Saccharomyces cerevisiae ), el nematodo Caenorhabditis elegans , la mosca común de la fruta ( Drosophila melanogaster ) y el ratón común ( Mus musculus ).
Medicina

La genética médica busca comprender cómo la variación genética se relaciona con la salud y la enfermedad humanas. Cuando se busca un gen desconocido que puede estar involucrado en una enfermedad, los investigadores utilizan comúnmente el enlace genético y los diagramas genéticos de pedigrí para encontrar la ubicación en el genoma asociado con la enfermedad. A nivel de la población, los investigadores aprovechan la aleatorización mendeliana para buscar ubicaciones en el genoma asociadas con enfermedades, un método especialmente útil para rasgos multigénicos no definidos claramente por un solo gen. Una vez que se encuentra un gen candidato, a menudo se realizan nuevas investigaciones sobre los genes correspondientes (u homólogos) de organismos modelo. Además de estudiar las enfermedades genéticas, la mayor disponibilidad de métodos de genotipado ha llevado al campo de la farmacogenética: el estudio de cómo el genotipo puede afectar las respuestas de los fármacos.
Las personas difieren en su tendencia hereditaria a desarrollar cáncer, y el cáncer es una enfermedad genética. El proceso de desarrollo del cáncer en el cuerpo es una combinación de eventos. Las mutaciones ocurren ocasionalmente dentro de las células del cuerpo a medida que se dividen. Aunque estas mutaciones no serán heredadas por ninguna descendencia, pueden afectar el comportamiento de las células, a veces causando que crezcan y se dividan con mayor frecuencia. Hay mecanismos biológicos que intentan detener este proceso; se dan señales a las células que se dividen de forma inapropiada que deberían desencadenar la muerte celular, pero a veces ocurren mutaciones adicionales que hacen que las células ignoren estos mensajes. Un proceso interno de selección natural ocurre dentro del cuerpo y eventualmente las mutaciones se acumulan dentro de las células para promover su propio crecimiento.
Normalmente, una célula se divide solo en respuesta a señales llamadas factores de crecimiento y deja de crecer una vez en contacto con las células circundantes y en respuesta a señales inhibidoras del crecimiento. Por lo general, se divide un número limitado de veces y muere, permaneciendo dentro del epitelio donde no puede migrar a otros órganos. Para convertirse en una célula cancerosa, una célula tiene que acumular mutaciones en varios genes (de tres a siete) que le permiten eludir esta regulación: ya no necesita dividir los factores de crecimiento, sigue creciendo al establecer contacto con las células vecinas, ignora las inhibidoras señales, sigue creciendo indefinidamente y es inmortal, escapa del epitelio y finalmente puede escapar del tumor primario, cruzar el endotelio de un vaso sanguíneo, ser transportado por el torrente sanguíneo y colonizar un nuevo órgano, formando una metástasis mortal. Aunque hay algunas predisposiciones genéticas en una pequeña fracción de cánceres, la fracción principal se debe a un conjunto de nuevas mutaciones genéticas que aparecen originalmente y se acumulan en una o un pequeño número de células que se dividirán para formar el tumor y no se transmiten a la progenie (mutaciones somáticas). Las mutaciones más frecuentes son la pérdida de la función de la proteína p53, un supresor tumoral o la vía p53 y la ganancia de mutaciones funcionales en las proteínas Ras o en otros oncogenes.
Métodos de busqueda
El ADN puede ser manipulado en el laboratorio. Las enzimas de restricción son enzimas de uso común que cortan el ADN en secuencias específicas, produciendo fragmentos predecibles de ADN. Los fragmentos de ADN se pueden visualizar mediante el uso de electroforesis en gel, que separa los fragmentos según su longitud.
El uso de enzimas de ligadura permite conectar fragmentos de ADN. Al unir ("ligar") fragmentos de ADN de diferentes fuentes, los investigadores pueden crear ADN recombinante, el ADN a menudo asociado con organismos genéticamente modificados. El ADN recombinante se usa comúnmente en el contexto de plásmidos: moléculas circulares cortas de ADN con algunos genes en ellas. En el proceso conocido como clonación molecular, los investigadores pueden amplificar los fragmentos de ADN insertando plásmidos en bacterias y luego cultivándolos en placas de agar (para aislar clones de células de bacterias, la "clonación" también puede referirse a los diversos medios para crear clones (" clonal ") organismos).
El ADN también se puede amplificar usando un procedimiento llamado reacción en cadena de la polimerasa (PCR). Mediante el uso de secuencias cortas específicas de ADN, la PCR puede aislar y amplificar exponencialmente una región de ADN dirigida. Debido a que puede amplificarse a partir de cantidades extremadamente pequeñas de ADN, la PCR también se usa a menudo para detectar la presencia de secuencias de ADN específicas.
Secuenciación de ADN y genómica
La secuenciación del ADN, una de las tecnologías más fundamentales desarrolladas para estudiar la genética, permite a los investigadores determinar la secuencia de nucleótidos en los fragmentos de ADN. La técnica de secuenciación por terminación de cadena, desarrollada en 1977 por un equipo dirigido por Frederick Sanger, todavía se usa de forma rutinaria para secuenciar fragmentos de ADN. Usando esta tecnología, los investigadores han podido estudiar las secuencias moleculares asociadas con muchas enfermedades humanas.
Como la secuenciación se ha vuelto menos costosa, los investigadores han secuenciado los genomas de muchos organismos mediante un proceso llamado ensamblaje del genoma, que utiliza herramientas computacionales para unir secuencias de muchos fragmentos diferentes. Estas tecnologías se usaron para secuenciar el genoma humano en el Proyecto del Genoma Humano completado en 2003. Las nuevas tecnologías de secuenciación de alto rendimiento reducen drásticamente el costo de la secuenciación del ADN, y muchos investigadores esperan reducir el costo de resecuenciación del genoma humano a mil dólares.
La secuenciación de próxima generación (o secuenciación de alto rendimiento) surgió debido a la demanda cada vez mayor de secuenciación de bajo costo. Estas tecnologías de secuenciación permiten la producción de potencialmente millones de secuencias al mismo tiempo. La gran cantidad de datos de secuencias disponibles ha creado el campo de la genómica, una investigación que utiliza herramientas computacionales para buscar y analizar patrones en los genomas completos de los organismos. La genómica también se puede considerar un subcampo de la bioinformática, que utiliza enfoques computacionales para analizar grandes conjuntos de datos biológicos. Un problema común en estos campos de investigación es cómo administrar y compartir datos que tratan temas humanos e información de identificación personal. Ver también compartir datos genómicos.
sociedad y Cultura
El 19 de marzo de 2015, un grupo de destacados biólogos instó a que se prohibiera en todo el mundo el uso clínico de métodos, en particular el uso de CRISPR y el dedo de zinc, para editar el genoma humano de una manera que pueda heredarse. En abril de 2015, investigadores chinos informaron resultados de investigaciones básicas para editar el ADN de embriones humanos no viables utilizando CRISPR.