Herencia
Definición
La herencia es la transmisión de los rasgos de los padres a sus descendientes, ya sea a través de la reproducción asexual o la reproducción sexual, las células descendientes u organismos adquieren la información genética de sus padres. A través de la herencia, las variaciones entre individuos pueden acumularse y hacer que las especies evolucionen por selección natural. El estudio de la herencia en biología es genética.Visión de conjunto


En los seres humanos, el color de los ojos es un ejemplo de una característica heredada: un individuo puede heredar el "rasgo de ojos marrones" de uno de los padres. Los rasgos heredados están controlados por genes y el conjunto completo de genes dentro del genoma de un organismo se llama genotipo.
El conjunto completo de rasgos observables de la estructura y el comportamiento de un organismo se llama fenotipo. Estos rasgos surgen de la interacción de su genotipo con el medio ambiente. Como resultado, muchos aspectos del fenotipo de un organismo no se heredan. Por ejemplo, la piel bronceada proviene de la interacción entre el fenotipo de una persona y la luz solar; por lo tanto, el bronceado no se transmite a los niños de las personas. Sin embargo, algunas personas se broncean más fácilmente que otras, debido a las diferencias en su genotipo: un ejemplo notable es la gente con el rasgo heredado de albinismo, que no se broncean y son muy sensibles a las quemaduras solares.
Se sabe que los rasgos heredables pasan de una generación a otra a través del ADN, una molécula que codifica información genética. El ADN es un polímero largo que incorpora cuatro tipos de bases, que son intercambiables. La secuencia de bases a lo largo de una molécula de ADN particular especifica la información genética: esto es comparable a una secuencia de letras que deletrea un pasaje de texto. Antes de que una célula se divida a través de la mitosis, el ADN se copia, de modo que cada una de las dos células resultantes heredará la secuencia de ADN. Una porción de una molécula de ADN que especifica una sola unidad funcional se llama gen; diferentes genes tienen diferentes secuencias de bases. Dentro de las células, las largas cadenas de ADN forman estructuras condensadas llamadas cromosomas. Los organismos heredan material genético de sus padres en forma de cromosomas homólogos, que contiene una combinación única de secuencias de ADN que codifican genes. La ubicación específica de una secuencia de ADN dentro de un cromosoma se conoce como un locus. Si la secuencia de ADN en un locus particular varía entre individuos, las diferentes formas de esta secuencia se denominan alelos. Las secuencias de ADN pueden cambiar a través de mutaciones, produciendo nuevos alelos. Si se produce una mutación dentro de un gen, el nuevo alelo puede afectar el rasgo que controla el gen, alterando el fenotipo del organismo.
Sin embargo, aunque esta simple correspondencia entre un alelo y un rasgo funciona en algunos casos, la mayoría de los rasgos son más complejos y están controlados por múltiples genes que interactúan dentro de los organismos y entre ellos. Los biólogos del desarrollo sugieren que las interacciones complejas en las redes genéticas y la comunicación entre las células pueden conducir a variaciones heredables que pueden ser la base de algunas de las mecánicas en la plasticidad y la canalización del desarrollo.
Los hallazgos recientes han confirmado ejemplos importantes de cambios hereditarios que no pueden explicarse por acción directa de la molécula de ADN. Estos fenómenos se clasifican como sistemas de herencia epigenéticos que evolucionan causalmente o independientemente sobre los genes. La investigación sobre modos y mecanismos de herencia epigenética aún está en su infancia científica, sin embargo, esta área de investigación ha atraído mucha actividad reciente ya que amplía el alcance de la heredabilidad y la biología evolutiva en general. La metilación del ADN que marca la cromatina, los bucles metabólicos autosostenidos, el silenciamiento génico por interferencia de ARN y la conformación tridimensional de las proteínas (como los priones) son áreas en las que se han descubierto sistemas de herencia epigenéticos a nivel organísmico. escamas. Por ejemplo, la herencia ecológica a través del proceso de construcción del nicho se define por las actividades regulares y repetidas de los organismos en su entorno. Esto genera un legado de efecto que modifica y retroalimenta el régimen de selección de las siguientes generaciones. Los descendientes heredan genes más las características ambientales generadas por las acciones ecológicas de los antepasados. Otros ejemplos de heredabilidad en la evolución que no están bajo el control directo de los genes incluyen la herencia de los rasgos culturales, la heredabilidad grupal y la simbiogénesis. Estos ejemplos de heredabilidad que operan por encima del gen se cubren ampliamente bajo el título de selección multinivel o jerárquica, que ha sido un tema de intenso debate en la historia de la ciencia evolutiva.
Relación con la teoría de la evolución
Cuando Charles Darwin propuso su teoría de la evolución en 1859, uno de sus principales problemas fue la falta de un mecanismo subyacente para la herencia. Darwin creía en una mezcla de herencia combinada y herencia de rasgos adquiridos (pangénesis). La combinación de la herencia conduciría a la uniformidad entre las poblaciones en solo unas pocas generaciones y luego eliminaría la variación de una población en la que podría actuar la selección natural. Esto llevó a Darwin a adoptar algunas ideas lamarckianas en ediciones posteriores de Sobre el origen de las especies y sus trabajos biológicos posteriores. El enfoque principal de Darwin hacia la herencia fue describir cómo parecía funcionar (darse cuenta de que los rasgos que no se expresaban explícitamente en el padre en el momento de la reproducción podían heredarse, que ciertos rasgos podían estar relacionados con el sexo, etc.) en lugar de sugerir mecanismos .
El modelo inicial de herencia de Darwin fue adoptado por su primo Francis Galton, y luego fuertemente modificado por él, quien estableció el marco para la escuela biométrica de la herencia. Galton no encontró evidencia para apoyar los aspectos del modelo de pangénesis de Darwin, que se basó en los rasgos adquiridos.
Se demostró que la herencia de los rasgos adquiridos tenía poca base en la década de 1880 cuando August Weismann cortó las colas de muchas generaciones de ratones y descubrió que su descendencia seguía desarrollando colas.
Historia

Los científicos en la antigüedad tenían una variedad de ideas sobre la herencia: Teofrasto propuso que las flores masculinas causaron que las flores femeninas maduraran, Hipócrates especuló que las "semillas" fueron producidas por varias partes del cuerpo y transmitidas a la descendencia en el momento de la concepción; y Aristóteles pensó que los fluidos masculinos y femeninos se mezclaron en la concepción. Esquilo, en 458 aC, propuso al macho como padre, con la hembra como "enfermera de la vida joven sembrada dentro de ella".
Las antiguas interpretaciones de la herencia hicieron la transición a dos doctrinas debatidas en el siglo XVIII. La Doctrina de la Epigenesia y la Doctrina de la Preformación fueron dos puntos de vista distintos sobre la comprensión de la herencia. La Doctrina de la epigénesis, originada por Aristóteles, afirmaba que un embrión se desarrolla continuamente. Las modificaciones de los rasgos del padre pasan a un embrión durante su vida. La base de esta doctrina se basa en la teoría de la herencia de los rasgos adquiridos. En oposición directa, la Doctrina de la Preformación afirmó que "genera lo mismo" donde el germen evolucionaría para producir descendientes similares a los padres. La visión preformacionista creía que la procreación era un acto de revelar lo que se había creado mucho antes. Sin embargo, esto fue disputado por la creación de la teoría celular en el siglo XIX, donde la unidad fundamental de la vida es la célula, y no algunas partes preformadas de un organismo. También se previeron varios mecanismos hereditarios, incluida la herencia de mezcla, sin ser probados o cuantificados adecuadamente, y luego fueron discutidos. Sin embargo, las personas pudieron desarrollar razas domésticas de animales y cultivos mediante selección artificial. La herencia de los rasgos adquiridos también formó parte de las primeras ideas lamarckianas sobre la evolución.
Durante el siglo XVIII, el microscopista holandés Antonie van Leeuwenhoek (1632-1723) descubrió "animálculos" en el esperma de humanos y otros animales. Algunos científicos especulan que vieron un "hombre pequeño" (homúnculo) dentro de cada esperma. Estos científicos formaron una escuela de pensamiento conocida como los "espermitas". Afirmaron que las únicas contribuciones de la hembra a la siguiente generación fueron el útero en el que creció el homúnculo y las influencias prenatales del útero. Una escuela opuesta de pensamiento, los ovists, creían que el futuro humano estaba en el huevo, y que el esperma simplemente estimulaba el crecimiento del huevo. Ovists pensó que las mujeres llevaban huevos que contenían niños y niñas, y que el sexo de la descendencia se determinó mucho antes de la concepción.
Gregor Mendel: padre de la genética

La idea de la herencia de partículas de genes se puede atribuir al monje de Moravia Gregor Mendel, quien publicó su trabajo sobre las plantas de guisantes en 1865. Sin embargo, su trabajo no fue ampliamente conocido y fue redescubierto en 1901. Inicialmente se asumió que la herencia mendeliana solo representaba diferencias grandes (cualitativas), como las observadas por Mendel en sus plantas de guisantes, y la idea de efecto aditivo de genes (cuantitativos) no se realizó hasta el trabajo de RA Fisher (1918), "La correlación entre parientes sobre la suposición de la herencia mendeliana "La contribución general de Mendel dio a los científicos una visión general útil de que los rasgos eran heredables. Su demostración de planta de guisantes se convirtió en la base del estudio de los Rasgos Mendelianos. Estos rasgos se pueden rastrear en un solo lugar.
Desarrollo moderno de genética y herencia
En la década de 1930, el trabajo de Fisher y otros resultó en una combinación de escuelas mendelianas y biométricas en la síntesis evolutiva moderna. La síntesis moderna cerró la brecha entre genetistas experimentales y naturalistas; y entre ambos y los paleontólogos, indicando que:
- Todos los fenómenos evolutivos pueden explicarse de manera consistente con los mecanismos genéticos conocidos y la evidencia observacional de los naturalistas.
- La evolución es gradual: pequeños cambios genéticos, recombinación ordenada por selección natural. Se explica que las discontinuidades entre especies (u otros taxones) se originan gradualmente a través de la separación geográfica y la extinción (no saltación).
- La selección es abrumadoramente el principal mecanismo de cambio; incluso pequeñas ventajas son importantes cuando se continúa. El objeto de la selección es el fenotipo en su ambiente circundante. El papel de la deriva genética es equívoco; aunque fue fuertemente respaldado inicialmente por Dobzhansky, fue degradado más tarde a medida que se obtenían los resultados de la genética ecológica.
- La primacía del pensamiento de la población: la diversidad genética que se transmite en las poblaciones naturales es un factor clave en la evolución. La fuerza de la selección natural en la naturaleza fue mayor de lo esperado; el efecto de factores ecológicos como la ocupación de nicho y la importancia de las barreras al flujo de genes son todos importantes.
La idea de que la especiación ocurre después de que las poblaciones se hayan aislado reproductivamente ha sido muy debatida. En las plantas, la poliploidía debe incluirse en cualquier vista de especiación. Las formulaciones tales como 'la evolución consiste principalmente en cambios en las frecuencias de los alelos entre una generación y otra' se propusieron bastante más tarde. La visión tradicional es que la biología del desarrollo ('evo-devo') jugó una pequeña parte en la síntesis, pero un relato de la obra de Gavin de Beer de Stephen Jay Gould sugiere que puede ser una excepción.
Casi todos los aspectos de la síntesis han sido desafiados a veces, con diversos grados de éxito. Sin embargo, no hay duda de que la síntesis fue un gran hito en la biología evolutiva. Aclaró muchas confusiones y fue directamente responsable de estimular una gran cantidad de investigación en la era posterior a la Segunda Guerra Mundial.
Trofim Lysenko, sin embargo, causó una reacción negativa de lo que ahora se llama Lysenkoism en la Unión Soviética cuando hizo hincapié en las ideas lamarckianas sobre la herencia de los rasgos adquiridos. Este movimiento afectó la investigación agrícola y condujo a la escasez de alimentos en la década de 1960 y afectó seriamente a la URSS.
Existe una evidencia creciente de que existe herencia transgeneracional de cambios epigenéticos en humanos y otros animales.
Desordenes genéticos comunes
- Síndrome de Down
- enfermedad de célula falciforme
- Fenilcetonuria (PKU)
- Hemofilia
Tipos
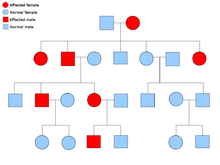
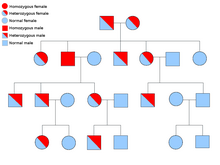
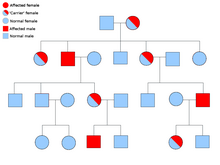
Alelos dominantes y recesivos
Se dice que un alelo es dominante si siempre se expresa en la apariencia de un organismo (fenotipo), siempre que esté presente al menos una copia del mismo. Por ejemplo, en los guisantes, el alelo para las vainas verdes, G , es dominante para las vainas amarillas, g . Por lo tanto, las plantas de guisantes con el par de alelos ya sea GG (homocigoto) o Gg (heterocigoto) tendrán vainas verdes. El alelo de las vainas amarillas es recesivo. Los efectos de este alelo solo se observan cuando está presente en ambos cromosomas, gg (homocigoto).
La descripción de un modo de herencia biológica consta de tres categorías principales:
- 1. Número de loci involucrados
- Monogenético (también llamado "simple") - un locus
- Oligogenético - pocos loci
- Poligenético - muchos loci
- 2. cromosomas implicados
- Autosómica - loci no están situados en un cromosoma sexual
- Gonosomal - loci están situados en un cromosoma sexual
- Los loci X-cromosómicos están situados en el cromosoma X (el caso más común)
- Los loci cromosómicos Y están situados en el cromosoma Y
- Mitocondrial: los loci están situados en el ADN mitocondrial
- 3. Correlación genotipo-fenotipo
- Dominante
- Intermedio (también llamado "codominante")
- Recesivo
- Sobredominante
- Underdominant
Estas tres categorías son parte de cada descripción exacta de un modo de herencia en el orden anterior. Además, se pueden agregar más especificaciones de la siguiente manera:
- 4. Interacciones casuales y ambientales
- Penetrancia
- Completar
- Incompleto (número porcentual)
- Expresividad
- Invariable
- Variable
- Heredabilidad (en modos de herencia poligénica y algunas veces también en oligogenética)
- Fenómenos de impresión materna o paterna (también ver epigenética)
- Penetrancia
- 5. Interacciones ligadas al sexo
- Herencia ligada al sexo (loci gonosomales)
- Expresión de fenotipo de sexo limitado (p. Ej., Criptorquismo)
- Herencia a través de la línea materna (en el caso de loci de ADN mitocondrial)
- Herencia a través de la línea paterna (en el caso de loci cromosómicos Y)
- 6. Interacciones locus-locus
- Epistasis con otros loci (por ejemplo, overdominance)
- Acoplamiento de genes con otros loci (también vea el cruce)
- Factores letales homocigotos
- Factores semi-letales
La determinación y descripción de un modo de herencia también se logra principalmente a través del análisis estadístico de los datos de pedigrí. En caso de que se conozcan los loci implicados, también se pueden emplear métodos de genética molecular.