RNA (ácido ribonucleico)
Definición

El ácido ribonucleico ( ARN ) es una molécula polimérica esencial en diversas funciones biológicas en la codificación, decodificación, regulación y expresión de genes. El ARN y el ADN son ácidos nucleicos y, junto con los lípidos, las proteínas y los carbohidratos, constituyen las cuatro macromoléculas principales esenciales para todas las formas de vida conocidas. Al igual que el ADN, el ARN se ensambla como una cadena de nucleótidos, pero a diferencia del ADN, se encuentra más a menudo en la naturaleza como una sola hebra plegada sobre sí misma, en lugar de una doble hebra apareada. Los organismos celulares usan ARN mensajero ( ARNm ) para transportar información genética (utilizando las bases nitrogenadas de guanina, uracilo, adenina y citosina, indicadas por las letras G, U, A y C) que dirigen la síntesis de proteínas específicas. Muchos virus codifican su información genética usando un genoma de ARN.
Algunas moléculas de ARN desempeñan un papel activo dentro de las células catalizando reacciones biológicas, controlando la expresión génica o detectando y comunicando respuestas a señales celulares. Uno de estos procesos activos es la síntesis de proteínas, una función universal en la que las moléculas de ARN dirigen el ensamblaje de proteínas en los ribosomas. Este proceso utiliza moléculas de ARN de transferencia ( ARNt ) para administrar aminoácidos al ribosoma, donde el ARN ribosomal ( ARNr ) luego une aminoácidos para formar proteínas.
Comparación con ADN

Al igual que el ADN, la mayoría de los ARN biológicamente activos, incluidos el ARNm, ARNt, ARNr, ARNsn y otros ARN no codificantes, contienen secuencias autocomplementarias que permiten que partes del ARN se plieguen y aparejen consigo mismas para formar dobles hélices. El análisis de estos ARN ha revelado que están altamente estructurados. A diferencia del ADN, sus estructuras no consisten en dobles hélices largas, sino más bien colecciones de hélices cortas agrupadas en estructuras similares a las proteínas. De esta manera, los ARN pueden lograr catálisis química (como enzimas). Por ejemplo, la determinación de la estructura del ribosoma-un complejo de ARN-proteína que cataliza la formación del enlace peptídico-reveló que su sitio activo está compuesto enteramente de ARN.
Estructura
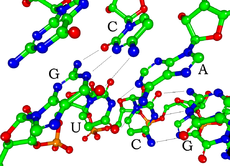
Cada nucleótido en el ARN contiene un azúcar de ribosa, con carbonos numerados del 1 'al 5'. Una base está unida a la posición 1 ', en general, adenina (A), citosina (C), guanina (G) o uracilo (U). La adenina y la guanina son purinas, la citosina y el uracilo son pirimidinas. Un grupo de fosfato se une a la posición 3 'de una ribosa y a la posición 5' de la siguiente. Los grupos fosfato tienen una carga negativa cada uno, lo que hace que el ARN sea una molécula cargada (polianión). Las bases forman enlaces de hidrógeno entre la citosina y la guanina, entre la adenina y el uracilo y entre la guanina y el uracilo. Sin embargo, otras interacciones son posibles, como un grupo de bases de adenina que se unen entre sí en un bulto, o el tetraloop de GNRA que tiene un par de bases guanina-adenina.

Un componente estructural importante del ARN que lo distingue del ADN es la presencia de un grupo hidroxilo en la posición 2 'del azúcar de ribosa. La presencia de este grupo funcional hace que la hélice tome principalmente la geometría de la forma A, aunque en contextos de dinucleótidos monocatenarios, el ARN raramente puede adoptar también la forma B más comúnmente observada en el ADN. La geometría de la forma A resulta en un surco mayor muy profundo y estrecho y un surco menor poco profundo y profundo. Una segunda consecuencia de la presencia del grupo 2'-hidroxilo es que en regiones conformacionalmente flexibles de una molécula de ARN (es decir, que no está involucrada en la formación de una doble hélice), puede atacar químicamente el enlace fosfodiéster adyacente para escindir la cadena principal.
El ARN se transcribe con solo cuatro bases (adenina, citosina, guanina y uracilo), pero estas bases y azúcares unidos se pueden modificar de numerosas maneras a medida que maduran los ARN. La pseudouridina (Ψ), en la que el enlace entre uracilo y ribosa se cambia de un enlace C-N a un enlace C-C, y ribotimidina (T) se encuentran en varios lugares (los más notables están en el bucle T ofC del tRNA ) Otra base modificada notable es la hipoxantina, una base de adenina desaminada cuyo nucleósido se llama inosina (I). La inosina desempeña un papel clave en la hipótesis de oscilación del código genético.
Hay más de 100 otros nucleósidos modificados naturales. La mayor diversidad estructural de modificaciones se puede encontrar en tRNA, mientras que pseudouridina y nucleósidos con 2'-O-methylribose a menudo presentes en rRNA son los más comunes. Las funciones específicas de muchas de estas modificaciones en el ARN no se comprenden completamente. Sin embargo, es notable que, en el ARN ribosomal, muchas de las modificaciones postranscripcionales ocurren en regiones altamente funcionales, como el centro peptidil transferasa y la interfaz de la subunidad, lo que implica que son importantes para la función normal.
La forma funcional de las moléculas de ARN monocatenarias, al igual que las proteínas, con frecuencia requiere una estructura terciaria específica. El andamio para esta estructura es proporcionado por elementos estructurales secundarios que son enlaces de hidrógeno dentro de la molécula. Esto conduce a varios "dominios" reconocibles de estructura secundaria, como bucles en horquilla, protuberancias y bucles internos. Como el ARN está cargado, se necesitan iones metálicos como el Mg para estabilizar muchas estructuras secundarias y terciarias.
El enantiómero natural del ARN es el ARN- D compuesto por D- ribonucleótidos. Todos los centros de quiralidad se encuentran en la D -ribosa. Mediante el uso de L -ribosa o más bien L-ribonucleótidos, se puede sintetizar L- ARN. El L- ARN es mucho más estable contra la degradación por RNasa.
Al igual que otros biopolímeros estructurados como las proteínas, se puede definir la topología de una molécula de ARN plegada. Esto se hace a menudo en base a la disposición de contactos dentro de la cadena dentro de un ARN plegado, denominado topología de circuito.
Síntesis
La síntesis de ARN generalmente es catalizada por una enzima ARN polimerasa que usa ADN como molde, un proceso conocido como transcripción. La iniciación de la transcripción comienza con la unión de la enzima a una secuencia promotora en el ADN (generalmente se encuentra "corriente arriba" de un gen). La doble hélice de ADN se desenrolla por la actividad helicasa de la enzima. La enzima luego progresa a lo largo de la cadena molde en la dirección 3 'a 5', sintetizando una molécula de ARN complementario con elongación que ocurre en la dirección 5 'a 3'. La secuencia de ADN también dicta dónde tendrá lugar la terminación de la síntesis de ARN.
Los ARN de transcripción primaria a menudo son modificados por enzimas después de la transcripción. Por ejemplo, se añaden una cola poli (A) y una tapa 5 'al pre-ARNm eucariótico y los intrones se eliminan mediante el spliceosoma.
También hay varias ARN polimerasas dependientes de ARN que utilizan ARN como plantilla para la síntesis de una nueva cadena de ARN. Por ejemplo, varios virus de ARN (como el poliovirus) usan este tipo de enzima para replicar su material genético. Además, la ARN polimerasa dependiente de ARN es parte de la ruta de interferencia de ARN en muchos organismos.
Tipos de ARN
Visión de conjunto

El ARN mensajero (ARNm) es el ARN que transporta información desde el ADN al ribosoma, los sitios de síntesis de proteínas (traducción) en la célula. La secuencia codificante del ARNm determina la secuencia de aminoácidos en la proteína que se produce. Sin embargo, muchos ARN no codifican proteínas (aproximadamente el 97% de la producción transcripcional no codifica proteínas en eucariotas).
Estos llamados ARN no codificantes ("ncRNA") pueden ser codificados por sus propios genes (genes de ARN), pero también pueden derivar de intrones de ARNm. Los ejemplos más destacados de ARN no codificantes son ARN de transferencia (ARNt) y ARN ribosómico (ARNr), que están implicados en el proceso de traducción. También hay ARN no codificantes implicados en la regulación de genes, procesamiento de ARN y otras funciones. Ciertos ARN son capaces de catalizar reacciones químicas tales como cortar y ligar otras moléculas de ARN, y la catálisis de la formación de enlaces peptídicos en el ribosoma; estos se conocen como ribozimas.
En longitud
De acuerdo con la longitud de la cadena de ARN, el ARN incluye ARN pequeño y ARN largo. Por lo general, los ARN pequeños tienen una longitud inferior a 200 nt, y los ARN largos tienen una longitud superior a 200 nt. Los ARN largos, también llamados ARN grandes, incluyen principalmente ARN largos no codificantes (ARN lnc) y ARNm. Los ARN pequeños incluyen principalmente ARN ribosómico 5.8S (ARNr), ARNr 5S, ARN de transferencia (ARNt), microARN (miARN), ARN interferente pequeño (ARNip), ARN nucleolar pequeño (ARNsn), ARN que interacciona con Piwi (ARNpi), ARNt- RNA pequeño derivado (tsRNA) y RNA derivado de rDNA pequeño (srRNA).
En traducción
El ARN mensajero (ARNm) transporta información sobre una secuencia de proteínas a los ribosomas, las fábricas de síntesis de proteínas en la célula. Está codificado de modo que cada tres nucleótidos (un codón) corresponde a un aminoácido. En células eucariotas, una vez que el ARNm precursor (pre-ARNm) se ha transcrito a partir del ADN, se procesa para que madure el ARNm. Esto elimina sus secciones intrones-no codificantes del pre-ARNm. El ARNm se exporta desde el núcleo al citoplasma, donde se une a los ribosomas y se traduce a su forma de proteína correspondiente con la ayuda del ARNt. En las células procariotas, que no tienen compartimentos de núcleo y citoplasma, el ARNm puede unirse a los ribosomas mientras se transcribe desde el ADN. Después de una cierta cantidad de tiempo, el mensaje se degrada en sus nucleótidos componentes con la ayuda de ribonucleasas.
El ARN de transferencia (ARNt) es una pequeña cadena de ARN de aproximadamente 80 nucleótidos que transfiere un aminoácido específico a una cadena polipeptídica en crecimiento en el sitio ribosómico de la síntesis de proteínas durante la traducción. Tiene sitios para la unión de aminoácidos y una región anticodón para el reconocimiento de codones que se une a una secuencia específica en la cadena de ARN mensajero a través de enlaces de hidrógeno.
El ARN ribosomal (ARNr) es el componente catalítico de los ribosomas. Los ribosomas eucarióticos contienen cuatro moléculas diferentes de rRNA: 18S, 5.8S, 28S y 5S rRNA. Tres de las moléculas de ARNr se sintetizan en el nucléolo y una se sintetiza en otro lugar. En el citoplasma, el ARN ribosomal y la proteína se combinan para formar una nucleoproteína llamada ribosoma. El ribosoma se une al ARNm y lleva a cabo la síntesis de proteínas. Varios ribosomas se pueden unir a un único ARNm en cualquier momento. Casi todo el ARN encontrado en una célula eucariota típica es ARNr.
El RNA mensajero de transferencia (tmRNA) se encuentra en muchas bacterias y plastidios. Etiqueta proteínas codificadas por mRNAs que carecen de codones de parada para la degradación e impide que el ribosoma se estanque.
ARN reguladores
Varios tipos de ARN pueden regular negativamente la expresión génica al ser complementarios a una parte de un ARNm o el ADN de un gen. Los microARN (miARN, 21-22 nt) se encuentran en eucariotas y actúan a través de interferencia de ARN (ARNi), donde un complejo efector de miARN y enzimas puede escindir ARNm complementario, bloquear el ARNm para su traducción o acelerar su degradación.
Mientras que los pequeños ARN interferentes (ARNip, 20-25 nt) a menudo se producen por la descomposición del ARN viral, también existen fuentes endógenas de ARNip. Los siRNAs actúan a través de la interferencia de RNA de una manera similar a miRNAs. Algunos miRNAs y siRNAs pueden provocar que los genes que se apuntan sean metilados, disminuyendo o aumentando la transcripción de esos genes. Los animales tienen RNA que interaccionan con Piwi (piRNA, 29-30 nt) que son activos en las células germinales y se cree que son una defensa contra los transposones y juegan un papel en la gametogénesis.
Muchos procariotas tienen ARN CRISPR, un sistema regulador similar a la interferencia de ARN. Los ARN antisentido están muy extendidos; la mayoría regula negativamente un gen, pero unos pocos son activadores de la transcripción. Una forma en que el ARN antisentido puede actuar es uniéndose a un ARNm, formando ARN bicatenario que se degrada enzimáticamente. Hay muchos ARN largos no codificantes que regulan genes en eucariotas, uno de los cuales es Xist, que recubre un cromosoma X en mamíferos hembras y lo inactiva.
Un ARNm puede contener elementos reguladores en sí mismo, tales como riboswitches, en la región 5 'no traducida o en la región 3' no traducida; estos elementos reguladores cis regulan la actividad de ese ARNm. Las regiones no traducidas también pueden contener elementos que regulan otros genes.
En procesamiento de ARN

Muchos ARN están involucrados en la modificación de otros ARN. Los intrones se empalman del pre-ARNm mediante los spliceosomas, que contienen varios ARN nucleares pequeños (snARN), o los intrones pueden ser ribozimas que se cortan solos. El ARN también puede alterarse haciendo que sus nucleótidos se modifiquen a nucleótidos distintos de A, C, G y U. En eucariotas, las modificaciones de los nucleótidos de ARN en general están dirigidas por pequeños ARN nucleolares (ARN sno; 60-300 nt), que se encuentran en el nucléolo y cuerpos de cajal. Los snoRNA se asocian con las enzimas y las guían a una mancha en un RNA emparejando a ese RNA. Estas enzimas luego realizan la modificación de nucleótidos. Los rRNA y los tRNA se modifican ampliamente, pero los snRNA y los mRNA también pueden ser el objetivo de la modificación de la base. El ARN también puede ser metilado.
Genomas de ARN
Al igual que el ADN, el ARN puede transportar información genética. Los virus de ARN tienen genomas compuestos de ARN que codifica una cantidad de proteínas. El genoma viral es replicado por algunas de esas proteínas, mientras que otras proteínas protegen el genoma a medida que la partícula del virus se traslada a una nueva célula huésped. Los viroides son otro grupo de patógenos, pero solo consisten en ARN, no codifican ninguna proteína y son replicados por la polimerasa de una célula vegetal hospedadora.
En transcripción inversa
Los virus de transcripción reversa replican sus genomas mediante la transcripción inversa de copias de ADN de su ARN; estas copias de ADN se transcriben a un nuevo ARN. Los retrotransposones también se propagan copiando ADN y ARN el uno del otro, y la telomerasa contiene un ARN que se usa como plantilla para construir los extremos de los cromosomas eucarióticos.
ARN bicatenario
El ARN bicatenario (ARNdc) es ARN con dos cadenas complementarias, similar al ADN que se encuentra en todas las células, pero con el reemplazo de la timina por el uracilo. dsRNA forma el material genético de algunos virus (virus de ARN bicatenario). El ARN bicatenario, como el ARN viral o ARNip, puede desencadenar interferencia de ARN en eucariotas, así como la respuesta al interferón en vertebrados.

ARN circular
A fines de la década de 1970, se demostró que existe una forma de ARN monocatenaria cerrada, es decir, circular, expresada en todo el reino animal y vegetal (ver circRNA). Se cree que los circRNAs surgen a través de una reacción de "back-splice" en la que el spliceosoma se une a un donador corriente abajo a un sitio de corte y empalme del aceptor aguas arriba. Hasta ahora, la función de los circRNAs es desconocida en gran medida, aunque en algunos ejemplos se ha demostrado una actividad de esponjado de microRNA.
Descubrimientos clave en la biología del ARN

La investigación sobre el ARN ha llevado a muchos descubrimientos biológicos importantes y numerosos premios Nobel. Los ácidos nucleicos fueron descubiertos en 1868 por Friedrich Miescher, quien llamó al material 'núcleo' desde que se encontró en el núcleo. Más tarde se descubrió que las células procariotas, que no tienen un núcleo, también contienen ácidos nucleicos. El papel del ARN en la síntesis de proteínas ya se sospechaba en 1939. Severo Ochoa ganó el Premio Nobel de Medicina en 1959 (compartido con Arthur Kornberg) después de que descubrió una enzima que puede sintetizar ARN en el laboratorio. Sin embargo, posteriormente se demostró que la enzima descubierta por Ochoa (polinucleótido fosforilasa) era responsable de la degradación del ARN, no de la síntesis del ARN.
La secuencia de los 77 nucleótidos de un ARNt de levadura fue encontrada por Robert W. Holley en 1965, ganando Holley el Premio Nobel de Medicina en 1968 (compartido con Har Gobind Khorana y Marshall Nirenberg).
Durante los primeros años de la década de 1970, se descubrieron retrovirus y transcriptasa inversa, demostrando por primera vez que las enzimas podían copiar el ARN en el ADN (lo contrario de la ruta habitual para la transmisión de información genética). Para este trabajo, David Baltimore, Renato Dulbecco y Howard Temin recibieron un Premio Nobel en 1975. En 1976, Walter Fiers y su equipo determinaron la primera secuencia completa de nucleótidos de un genoma del virus ARN, el del bacteriófago MS2.
En 1977, se descubrieron intrones y empalme de ARN tanto en virus de mamíferos como en genes celulares, lo que resultó en un Nobel de 1993 para Philip Sharp y Richard Roberts. Las moléculas de ARN catalítico (ribozimas) se descubrieron a principios de la década de 1980, lo que condujo a un premio Nobel de 1989 a Thomas Cech y Sidney Altman. En 1990, se encontró en Petunia que los genes introducidos pueden silenciar genes similares propios de la planta, que ahora se sabe que son el resultado de la interferencia del ARN.
Aproximadamente al mismo tiempo, se descubrió que los ARN de 22 nt de longitud, ahora llamados microARN, tienen un papel en el desarrollo de C. elegans . Los estudios sobre la interferencia de ARN obtuvieron un Premio Nobel para Andrew Fire y Craig Mello en 2006, y otro Nobel fue otorgado para estudios sobre la transcripción de ARN a Roger Kornberg en el mismo año. El descubrimiento de ARN reguladores de genes ha conducido a intentos de desarrollar fármacos hechos de ARN, como ARNip, para silenciar genes.
Relevancia para la química prebiótica y la abiogénesis
En 1967, Carl Woese planteó la hipótesis de que el ARN podría ser catalítico y sugirió que las formas de vida más antiguas (moléculas autorreplicantes) podrían haber dependido del ARN tanto para transportar información genética como para catalizar reacciones bioquímicas: un mundo de ARN.
En marzo de 2015, se informaron nucleótidos complejos de ADN y ARN, incluyendo uracilo, citosina y timina, en el laboratorio en condiciones de espacio exterior, utilizando productos químicos iniciadores, como la pirimidina, un compuesto orgánico que se encuentra comúnmente en los meteoritos. La pirimidina, al igual que los hidrocarburos aromáticos policíclicos (HAP), es uno de los compuestos más ricos en carbono que se encuentran en el Universo y puede haberse formado en gigantes rojos o en polvo interestelar y nubes de gas.