Sistema inmune
Definición
El sistema inmune es un sistema de defensa del huésped que comprende muchas estructuras biológicas y procesos dentro de un organismo que protege contra la enfermedad. Para funcionar correctamente, un sistema inmunitario debe detectar una amplia variedad de agentes, conocidos como patógenos, desde virus hasta gusanos parásitos, y distinguirlos del propio tejido sano del organismo. En muchas especies, el sistema inmune puede clasificarse en subsistemas, como el sistema inmune innato frente al sistema inmune adaptativo, o la inmunidad humoral frente a la inmunidad mediada por células. En los humanos, la barrera hematoencefálica, la barrera sangre-líquido cefalorraquídeo y las barreras cerebrales fluidas similares separan el sistema inmunitario periférico del sistema neuroinmune, que protege el cerebro.
Los patógenos pueden evolucionar y adaptarse rápidamente, y así evitar la detección y neutralización por parte del sistema inmune; sin embargo, también han evolucionado múltiples mecanismos de defensa para reconocer y neutralizar patógenos. Incluso organismos unicelulares simples como las bacterias poseen un sistema inmune rudimentario en forma de enzimas que protegen contra las infecciones por bacteriófagos. Otros mecanismos inmunes básicos evolucionaron en eucariotas antiguos y permanecen en sus descendientes modernos, como plantas e invertebrados. Estos mecanismos incluyen fagocitosis, péptidos antimicrobianos llamados defensinas y el sistema del complemento. Los vertebrados machacados, incluidos los humanos, tienen mecanismos de defensa aún más sofisticados, incluida la capacidad de adaptarse con el tiempo para reconocer patógenos específicos de manera más eficiente. La inmunidad adaptativa (o adquirida) crea memoria inmunológica después de una respuesta inicial a un patógeno específico, lo que lleva a una respuesta mejorada a los encuentros posteriores con ese mismo patógeno. Este proceso de inmunidad adquirida es la base de la vacunación.
Los desórdenes del sistema inmune pueden resultar en enfermedades autoinmunes, enfermedades inflamatorias y cáncer. La inmunodeficiencia se produce cuando el sistema inmunitario está menos activo de lo normal, lo que provoca infecciones recurrentes y potencialmente mortales. En los humanos, la inmunodeficiencia puede ser el resultado de una enfermedad genética, como la inmunodeficiencia combinada grave, afecciones adquiridas como el VIH / SIDA o el uso de medicamentos inmunosupresores. Por el contrario, la autoinmunidad es el resultado de un sistema inmune hiperactivo que ataca los tejidos normales como si fueran organismos extraños. Las enfermedades autoinmunes comunes incluyen tiroiditis de Hashimoto, artritis reumatoide, diabetes mellitus tipo 1 y lupus eritematoso sistémico. La inmunología cubre el estudio de todos los aspectos del sistema inmune.
Historia de la inmunología
La inmunología es una ciencia que examina la estructura y la función del sistema inmune. Se origina de la medicina y los primeros estudios sobre las causas de la inmunidad a la enfermedad. La referencia más antigua conocida a la inmunidad fue durante la plaga de Atenas en 430 aC Tucídides notó que las personas que se habían recuperado de una enfermedad anterior podían amamantar a los enfermos sin contraer la enfermedad por segunda vez. En el siglo XVIII, Pierre-Louis Moreau de Maupertuis hizo experimentos con veneno de escorpión y observó que ciertos perros y ratones eran inmunes a este veneno. Esta y otras observaciones de la inmunidad adquirida fueron explotadas más tarde por Louis Pasteur en su desarrollo de la vacunación y su teoría germinal propuesta de la enfermedad. La teoría del aficionado estaba en oposición directa a las teorías contemporáneas de la enfermedad, como la teoría del miasma. No fue hasta las pruebas de 1891 de Robert Koch, por las que se le concedió un Premio Nobel en 1905, que se confirmaron los microorganismos como la causa de las enfermedades infecciosas. Los virus fueron confirmados como patógenos humanos en 1901, con el descubrimiento del virus de la fiebre amarilla por Walter Reed.
La inmunología hizo un gran avance hacia finales del siglo XIX, a través de rápidos desarrollos, en el estudio de la inmunidad humoral y la inmunidad celular. Particularmente importante fue el trabajo de Paul Ehrlich, quien propuso la teoría de la cadena lateral para explicar la especificidad de la reacción antígeno-anticuerpo; sus contribuciones al entendimiento de la inmunidad humoral fueron reconocidas por la concesión de un Premio Nobel en 1908, que fue otorgado conjuntamente al fundador de la inmunología celular, Elie Metchnikoff.
Defensa en capas
El sistema inmune protege a los organismos de la infección con defensas estratificadas de especificidad creciente. En términos simples, las barreras físicas impiden que patógenos como bacterias y virus ingresen al organismo. Si un patógeno rompe estas barreras, el sistema inmune innato proporciona una respuesta inmediata, pero no específica. Los sistemas inmunes innatos se encuentran en todas las plantas y animales. Si los patógenos evaden con éxito la respuesta innata, los vertebrados poseen una segunda capa de protección, el sistema inmune adaptativo, que se activa mediante la respuesta innata. Aquí, el sistema inmune adapta su respuesta durante una infección para mejorar su reconocimiento del patógeno. Esta respuesta mejorada se retiene luego de eliminar el patógeno, en forma de memoria inmunológica.
| Sistema inmune innato | Sistema inmune adaptativo |
|---|---|
| La respuesta no es específica | Respuesta específica de patógeno y antígeno |
| La exposición conduce a una respuesta máxima inmediata | Tiempo de retardo entre la exposición y la respuesta máxima |
| Componentes humorales y mediados por células | Componentes humorales y mediados por células |
| Sin memoria inmunológica | La exposición conduce a la memoria inmunológica |
| Encontrado en casi todas las formas de vida | Se encuentra solo en vertebrados con mandíbulas |
Tanto la inmunidad innata como la adaptativa dependen de la capacidad del sistema inmune para distinguir entre moléculas propias y no propias. En inmunología, las auto moléculas son aquellos componentes del cuerpo de un organismo que el sistema inmunitario puede distinguir de las sustancias extrañas. Por el contrario, las moléculas no propias son aquellas reconocidas como moléculas extrañas. Una clase de moléculas no propios se llaman antígenos (abreviatura de contracuerpo Gen erators) y se definen como sustancias que se unen a receptores inmunes específicas y provocan una respuesta inmune.
Sistema inmune innato
Los microorganismos o toxinas que entran con éxito en un organismo se encuentran con las células y los mecanismos del sistema inmune innato. La respuesta innata generalmente se desencadena cuando los microbios se identifican mediante receptores de reconocimiento de patrones, que reconocen componentes que se conservan entre amplios grupos de microorganismos, o cuando las células lesionadas, dañadas o estresadas envían señales de alarma, muchas de las cuales (pero no todas) por los mismos receptores que aquellos que reconocen patógenos. Las defensas innatas innatas no son específicas, lo que significa que estos sistemas responden a los patógenos de forma genérica. Este sistema no confiere inmunidad de larga duración contra un patógeno. El sistema inmune innato es el sistema dominante de defensa del huésped en la mayoría de los organismos.
Barreras de superficie
Varias barreras protegen a los organismos de las infecciones, incluidas las barreras mecánicas, químicas y biológicas. La cutícula cerosa de la mayoría de las hojas, el exoesqueleto de los insectos, las cáscaras y las membranas de los huevos depositados externamente y la piel son ejemplos de barreras mecánicas que son la primera línea de defensa contra la infección. Sin embargo, como los organismos no pueden ser completamente sellados de su entorno, otros sistemas actúan para proteger las aberturas del cuerpo, como los pulmones, los intestinos y el tracto genitourinario. En los pulmones, la tos y los estornudos expulsan mecánicamente los patógenos y otros irritantes del tracto respiratorio. La acción de enjuague de las lágrimas y la orina también expulsa mecánicamente a los patógenos, mientras que el moco secretado por el tracto respiratorio y gastrointestinal sirve para atrapar y enredar microorganismos.
Las barreras químicas también protegen contra la infección. La piel y el tracto respiratorio secretan péptidos antimicrobianos como las β-defensinas. Las enzimas como la lisozima y la fosfolipasa A2 en la saliva, las lágrimas y la leche materna también son antibacterianas. Las secreciones vaginales sirven como una barrera química después de la menarca, cuando se vuelven ligeramente ácidas, mientras que el semen contiene defensinas y zinc para matar a los patógenos. En el estómago, el ácido gástrico y las proteasas sirven como poderosas defensas químicas contra los patógenos ingeridos.
Dentro de los tractos genitourinario y gastrointestinal, la flora comensal sirve como barreras biológicas compitiendo con bacterias patógenas por alimentos y espacio y, en algunos casos, cambiando las condiciones en su entorno, como el pH o el hierro disponible. Como resultado de la relación simbiótica entre los comensales y el sistema inmune, se reduce la probabilidad de que los patógenos alcancen un número suficiente como para causar enfermedades. Sin embargo, dado que la mayoría de los antibióticos no se dirigen específicamente a las bacterias y no afectan a los hongos, los antibióticos orales pueden conducir a un "crecimiento excesivo" de hongos y causar condiciones como candidiasis vaginal (infección por hongos). Hay buena evidencia de que la reintroducción de la flora probiótica, como los cultivos puros de lactobacilos que normalmente se encuentran en el yogur no pasteurizado,
Inflamación
La inflamación es una de las primeras respuestas del sistema inmune a la infección. Los síntomas de la inflamación son enrojecimiento, hinchazón, calor y dolor, que son causados por un mayor flujo de sangre al tejido. La inflamación es producida por eicosanoides y citocinas, que son liberadas por las células lesionadas o infectadas. Los eicosanoides incluyen las prostaglandinas que producen fiebre y la dilatación de los vasos sanguíneos asociados con la inflamación, y los leucotrienos que atraen a ciertos glóbulos blancos (leucocitos). Las citocinas comunes incluyen interleuquinas que son responsables de la comunicación entre los glóbulos blancos; quimiocinas que promueven la quimiotaxis; e interferones que tienen efectos antivirales, como el cierre de la síntesis de proteínas en la célula huésped. También pueden liberarse factores de crecimiento y factores citotóxicos.
Sistema complementario
El sistema de complemento es una cascada bioquímica que ataca las superficies de células extrañas. Contiene más de 20 proteínas diferentes y se llama así por su capacidad de "complementar" la muerte de patógenos por anticuerpos. El complemento es el principal componente humoral de la respuesta inmune innata. Muchas especies tienen sistemas de complemento, incluidos mamíferos como plantas, peces y algunos invertebrados.
En los seres humanos, esta respuesta se activa por la unión del complemento a los anticuerpos que se han unido a estos microbios o la unión de las proteínas del complemento a los carbohidratos en las superficies de los microbios. Esta señal de reconocimiento desencadena una respuesta de muerte rápida. La velocidad de la respuesta es el resultado de la amplificación de la señal que ocurre después de la activación proteolítica secuencial de las moléculas del complemento, que también son proteasas. Después de que las proteínas del complemento se unen inicialmente al microbio, activan su actividad de proteasa, que a su vez activa otras proteasas del complemento, y así sucesivamente. Esto produce una cascada catalítica que amplifica la señal inicial mediante retroalimentación positiva controlada. La cascada da como resultado la producción de péptidos que atraen células inmunes, aumentan la permeabilidad vascular y opsonizan (recubren) la superficie de un patógeno, marcándolo para la destrucción. Esta deposición de complemento también puede matar células directamente al alterar su membrana plasmática.
Barreras celulares

Los leucocitos (glóbulos blancos) actúan como organismos independientes, unicelulares y son el segundo brazo del sistema inmune innato. Los leucocitos innatos incluyen los fagocitos (macrófagos, neutrófilos y células dendríticas), células linfoides innatas, mastocitos, eosinófilos, basófilos y células asesinas naturales. Estas células identifican y eliminan los patógenos, ya sea atacando a los patógenos más grandes a través del contacto o absorbiendo y luego matando a los microorganismos. Las células innatas también son importantes mediadores en el desarrollo de órganos linfoides y la activación del sistema inmune adaptativo.
La fagocitosis es una característica importante de la inmunidad celular innata realizada por células llamadas fagocitos que engullen o comen patógenos o partículas. Los fagocitos generalmente patrullan el cuerpo en busca de patógenos, pero las citocinas pueden llamarlo a lugares específicos. Una vez que un patógeno ha sido engullido por un fagocito, que queda atrapado en una vesícula intracelular llamada fagosoma, que posteriormente se fusiona con otra vesícula llamada lisosoma para formar un fagolisosoma. El patógeno es destruido por la actividad de las enzimas digestivas o después de una explosión respiratoria que libera radicales libres en el fagolisosoma. La fagocitosis evolucionó como un medio de adquisición de nutrientes, pero este papel se extendió en los fagocitos para incluir el engullimiento de los patógenos como mecanismo de defensa. La fagocitosis probablemente represente la forma más antigua de defensa del huésped,
Los neutrófilos y los macrófagos son fagocitos que viajan por todo el cuerpo en busca de patógenos invasores. Los neutrófilos se encuentran normalmente en el torrente sanguíneo y son el tipo más abundante de fagocitos, que normalmente representan del 50% al 60% del total de leucocitos circulantes, y consisten en subpoblaciones de neutrófilos y neutrófilos. Durante la fase aguda de la inflamación, particularmente como resultado de una infección bacteriana, los neutrófilos migran hacia el sitio de la inflamación en un proceso llamado quimiotaxis, y generalmente son las primeras células que llegan a la escena de la infección. Los macrófagos son células versátiles que residen en los tejidos y producen una amplia gama de sustancias químicas, incluidas enzimas, proteínas del complemento y citocinas, y también pueden actuar como carroñeros que liberan al cuerpo de las células gastadas y otros desechos.
Las células dendríticas (DC) son fagocitos en tejidos que están en contacto con el entorno externo; por lo tanto, se encuentran principalmente en la piel, la nariz, los pulmones, el estómago y los intestinos. Se nombran por su parecido con las dendritas neuronales, ya que ambos tienen muchas proyecciones similares a la columna vertebral, pero las células dendríticas no están conectadas de ninguna manera con el sistema nervioso. Las células dendríticas sirven como un vínculo entre los tejidos corporales y los sistemas inmune innato y adaptativo, ya que presentan antígenos a las células T, uno de los tipos de células clave del sistema inmune adaptativo.
Los mastocitos residen en los tejidos conectivos y las membranas mucosas, y regulan la respuesta inflamatoria. En la mayoría de los casos se asocian con alergia y anafilaxia. Los basófilos y los eosinófilos están relacionados con los neutrófilos. Secretan mediadores químicos que participan en la defensa contra parásitos y juegan un papel en las reacciones alérgicas, como el asma. Las células asesinas naturales (células NK) son leucocitos que atacan y destruyen células tumorales o células que han sido infectadas por virus.
Células asesinas naturales
Las células asesinas naturales, o células NK, son linfocitos y un componente del sistema inmune innato que no ataca directamente a los microbios invasores. Por el contrario, las células NK destruyen las células huésped comprometidas, como las células tumorales o las células infectadas por virus, reconociendo dichas células por una condición conocida como "falta de uno mismo". Este término describe células con bajos niveles de un marcador de superficie celular llamado MHC I (complejo de histocompatibilidad principal), una situación que puede surgir en infecciones virales de células hospedadoras. Fueron llamados "asesinos naturales" debido a la noción inicial de que no requieren activación para matar las células que se "pierden". Durante muchos años no estaba claro cómo las células NK reconocen las células tumorales y las células infectadas. Ahora se sabe que el maquillaje MHC en la superficie de esas células se altera y las células NK se activan a través del reconocimiento de "falta de uno mismo". Las células NK no reconocen y atacan las células corporales normales porque expresan antígenos de MHC auto intactos. Esos antígenos MHC son reconocidos por receptores de inmunoglobulina de células asesinas (KIR) que esencialmente ponen freno a las células NK.
Sistema inmune adaptativo
El sistema inmune adaptativo evolucionó en los primeros vertebrados y permite una respuesta inmune más fuerte, así como la memoria inmunológica, donde cada patógeno es "recordado" por un antígeno de firma. La respuesta inmune adaptativa es específica de antígeno y requiere el reconocimiento de antígenos específicos "no propios" durante un proceso denominado presentación de antígeno. La especificidad del antígeno permite la generación de respuestas que se adaptan a patógenos específicos o células infectadas por patógenos. La capacidad de montar estas respuestas adaptadas se mantiene en el cuerpo mediante "celdas de memoria". Si un patógeno infecta al cuerpo más de una vez, estas células de memoria específicas se utilizan para eliminarlo rápidamente.
Linfocitos
Las células del sistema inmune adaptativo son tipos especiales de leucocitos, llamados linfocitos. Las células B y las células T son los tipos principales de linfocitos y se derivan de células madre hematopoyéticas en la médula ósea. Las células B están involucradas en la respuesta inmune humoral, mientras que las células T están involucradas en la respuesta inmune mediada por células.
Tanto las células B como las células T transportan moléculas receptoras que reconocen objetivos específicos. Las células T reconocen un objetivo "no propio", como un patógeno, solo después de que los antígenos (pequeños fragmentos del patógeno) han sido procesados y presentados en combinación con un receptor "propio" llamado molécula del complejo mayor de histocompatibilidad (MHC). Hay dos subtipos principales de células T: el asesino T celland, el linfocito T auxiliar. Además, hay células T reguladoras que tienen un papel en la modulación de la respuesta inmune. Las células T asesinas solo reconocen antígenos acoplados a moléculas MHC de Clase I, mientras que las células T auxiliares y las células T reguladoras solo reconocen antígenos acoplados a moléculas MHC de Clase II. Estos dos mecanismos de presentación de antígenos reflejan los diferentes roles de los dos tipos de células T. Un tercio, subtipo menor son las células T γδ que reconocen antígenos intactos que no están unidos a los receptores MHC. Las células T doblemente positivas están expuestas a una amplia variedad de autoantígenos en el timo, en los que el yodo es necesario para su desarrollo y actividad del timo.
En contraste, el receptor específico de antígeno de célula B es una molécula de anticuerpo en la superficie de célula B, y reconoce patógenos completos sin necesidad de procesamiento de antígeno. Cada linaje de células B expresa un anticuerpo diferente, por lo que el conjunto completo de receptores de antígenos de células B representa todos los anticuerpos que el cuerpo puede fabricar.
Células T asesinas
Las células T asesinas son un subgrupo de células T que matan las células que están infectadas con virus (y otros patógenos), o están dañadas o son disfuncionales. Al igual que con las células B, cada tipo de célula T reconoce un antígeno diferente. Las células T asesinas se activan cuando su receptor de células T (TCR) se une a este antígeno específico en un complejo con el receptor MHC Clase I de otra célula. El reconocimiento de este MHC: complejo de antígeno es ayudado por un correceptor en la célula T, llamado CD8. La célula T luego viaja por todo el cuerpo en busca de células en las que los receptores MHC I portan este antígeno. Cuando una célula T activada entra en contacto con tales células, libera citotoxinas, como la perforina, que forma poros en la membrana plasmática de la célula diana, lo que permite la entrada de iones, agua y toxinas. La entrada de otra toxina llamada granulisina (una proteasa) induce a la célula diana a sufrir apoptosis. La destrucción de las células huésped por células T es particularmente importante para prevenir la replicación de virus. La activación de células T está estrechamente controlada y generalmente requiere una señal de activación de MHC / antígeno muy fuerte, o señales de activación adicionales proporcionadas por células T "auxiliares" (véase a continuación).
Helper T cells

Las células T auxiliares regulan tanto la respuesta inmune innata como la adaptativa y ayudan a determinar qué respuestas inmunitarias produce el cuerpo a un patógeno en particular. Estas células no tienen actividad citotóxica y no matan las células infectadas ni eliminan los patógenos directamente. En su lugar, controlan la respuesta inmune al dirigir a otras células para que realicen estas tareas.
Las células T colaboradoras expresan receptores de células T (TCR) que reconocen el antígeno unido a las moléculas MHC de Clase II. El complejo MHC: antígeno también es reconocido por el correceptor CD4 de la célula auxiliar, que recluta moléculas dentro de la célula T (por ejemplo, Lck) que son responsables de la activación de las células T. Las células T colaboradoras tienen una asociación más débil con el complejo MHC: antígeno que las células T asesinas, lo que significa que muchos receptores (alrededor de 200-300) en la célula T auxiliar deben estar unidos por un antígeno MHC: para activar la célula auxiliar. mientras que las células T asesinas se pueden activar mediante el acoplamiento de una única molécula MHC: antígeno. La activación de las células T colaboradoras también requiere una mayor duración de interacción con una célula presentadora de antígeno. La activación de una célula T auxiliar en reposo hace que libere citoquinas que influyen en la actividad de muchos tipos de células. Las señales de citocinas producidas por las células T auxiliares mejoran la función microbicida de los macrófagos y la actividad de las células T asesinas. Además, la activación de las células T auxiliares causa una regulación positiva de las moléculas expresadas en la superficie de las células T, como el ligando CD40 (también llamado CD154), que proporciona señales estimuladoras adicionales normalmente requeridas para activar las células B productoras de anticuerpos.
Gamma delta T cells
Gamma delta T cells (células T γδ) poseen un receptor alternativo de células T (TCR) en oposición a las células T CD4 + y CD8 + (αβ) y comparten las características de las células T auxiliares, las células T citotóxicas y las células NK. Las condiciones que producen respuestas de las células T γδ no se comprenden completamente. Al igual que otros subconjuntos de células T "no convencionales" que llevan TCR invariantes, como las células T asesinas naturales restringidas por CD1d, las células T γδ se encuentran en el límite entre la inmunidad innata y la adaptativa. Por un lado, las células T γδ son un componente de la inmunidad adaptativa ya que reorganizan los genes TCR para producir diversidad de receptores y también pueden desarrollar un fenotipo de memoria. Por otro lado, los diversos subconjuntos también son parte del sistema inmune innato, ya que los receptores restringidos de TCR o NK pueden usarse como receptores de reconocimiento de patrones. Por ejemplo,

Linfocitos B y anticuerpos
Una célula B identifica patógenos cuando los anticuerpos en su superficie se unen a un antígeno extraño específico. Este complejo antígeno / anticuerpo es absorbido por la célula B y procesado por proteólisis en péptidos. Luego, la célula B muestra estos péptidos antigénicos en su superficie moléculas MHC de clase II. Esta combinación de MHC y antígeno atrae a una célula T colaboradora coincidente, que libera linfocinas y activa la célula B. A medida que la célula B activada comienza a dividirse, su descendencia (células plasmáticas) secreta millones de copias del anticuerpo que reconoce este antígeno. Estos anticuerpos circulan en el plasma sanguíneo y la linfa, se unen a los patógenos que expresan el antígeno y los marcan para destrucción por activación del complemento o por captación y destrucción por fagocitos. Los anticuerpos también pueden neutralizar los desafíos directamente,
Sistema inmune adaptativo alternativo
La evolución del sistema inmune adaptativo ocurrió en un ancestro de los vertebrados con mandíbula. Muchas de las moléculas clásicas del sistema inmune adaptativo (p. Ej., Inmunoglobulinas y receptores de células T) existen solo en vertebrados con mandíbula. Sin embargo, se ha descubierto una molécula distinta derivada de linfocitos en vertebrados sin mandíbula primitivos, como la lamprea y el hagfish. Estos animales poseen una gran variedad de moléculas llamadas receptores de linfocitos variables (VLR) que, al igual que los receptores de antígenos de los vertebrados con mandíbula, se producen a partir de solo un pequeño número (uno o dos) de genes. Se cree que estas moléculas se unen a antígenos patógenos de forma similar a los anticuerpos, y con el mismo grado de especificidad.
Memoria inmunológica
Cuando las células B y las células T se activan y comienzan a replicarse, algunas de sus crías se convierten en células de memoria de larga vida. A lo largo de la vida de un animal, estas células de memoria recuerdan cada patógeno específico encontrado y pueden generar una respuesta fuerte si el patógeno se detecta nuevamente. Esto es "adaptativo" porque ocurre durante la vida de un individuo como una adaptación a la infección con ese patógeno y prepara al sistema inmune para los desafíos futuros. La memoria inmunológica puede ser en forma de memoria pasiva a corto plazo o memoria activa a largo plazo.
Memoria pasiva
Los recién nacidos no tienen exposición previa a los microbios y son particularmente vulnerables a las infecciones. La madre proporciona varias capas de protección pasiva. Durante el embarazo, un tipo particular de anticuerpo, llamado IgG, se transporta de la madre al bebé directamente a través de la placenta, por lo que los bebés humanos tienen altos niveles de anticuerpos incluso al nacer, con el mismo rango de especificidades antigénicas que su madre. La leche materna o el calostro también contienen anticuerpos que se transfieren al intestino del bebé y protegen contra las infecciones bacterianas hasta que el recién nacido pueda sintetizar sus propios anticuerpos. Esta es la inmunidad pasiva porque el feto en realidad no produce células de memoria o anticuerpos, solo los toma prestados. Esta inmunidad pasiva suele ser a corto plazo, con una duración de algunos días hasta varios meses. En medicina,

Memoria activa e inmunización
La memoria activa a largo plazo se adquiere después de la infección mediante la activación de las células B y T. La inmunidad activa también se puede generar artificialmente, a través de la vacunación. El principio detrás de la vacunación (también llamada inmunización) es introducir un antígeno de un patógeno para estimular el sistema inmune y desarrollar inmunidad específica contra ese patógeno en particular sin causar enfermedad asociada con ese organismo. Esta inducción deliberada de una respuesta inmune es exitosa porque explota la especificidad natural del sistema inmune, así como su inducibilidad. Con las enfermedades infecciosas siendo una de las principales causas de muerte en la población humana, la vacunación representa la manipulación más efectiva del sistema inmune que la humanidad ha desarrollado.
La mayoría de las vacunas virales se basan en virus vivos atenuados, mientras que muchas vacunas bacterianas se basan en componentes acelulares de microorganismos, incluidos los componentes tóxicos inocuos. Dado que muchos antígenos derivados de vacunas acelulares no inducen fuertemente la respuesta adaptativa, la mayoría de las vacunas bacterianas se proporcionan con adyuvantes adicionales que activan las células presentadoras de antígeno del sistema inmune innato y maximizan la inmunogenicidad.
Trastornos de la inmunidad humana
El sistema inmune es una estructura notablemente efectiva que incorpora especificidad, inducibilidad y adaptación. Sin embargo, se producen fallas en la defensa del huésped y se dividen en tres grandes categorías: inmunodeficiencias, autoinmunidad e hipersensibilidades.
Inmunodeficiencias
Las inmunodeficiencias ocurren cuando uno o más de los componentes del sistema inmune están inactivos. La capacidad del sistema inmune para responder a los patógenos disminuye tanto en los jóvenes como en los ancianos, y las respuestas inmunes comienzan a disminuir alrededor de los 50 años de edad debido a la inmunosenescencia. En los países desarrollados, la obesidad, el alcoholismo y el consumo de drogas son causas comunes de una función inmune pobre. Sin embargo, la desnutrición es la causa más común de inmunodeficiencia en los países en desarrollo. Las dietas que carecen de suficiente proteína se asocian con la inmunidad mediada por células, la actividad del complemento, la función de los fagocitos, las concentraciones de anticuerpos IgA y la producción de citocinas. Además, la pérdida del timo a una edad temprana a través de la mutación genética o la extirpación quirúrgica da como resultado una inmunodeficiencia grave y una alta susceptibilidad a la infección.
Las inmunodeficiencias también pueden ser heredadas o 'adquiridas'. La enfermedad granulomatosa crónica, donde los fagocitos tienen una capacidad reducida para destruir los patógenos, es un ejemplo de una inmunodeficiencia hereditaria o congénita. El SIDA y algunos tipos de cáncer causan inmunodeficiencia adquirida.
Autoinmunidad
Las respuestas inmunitarias hiperactivas comprenden el otro extremo de la disfunción inmune, particularmente los trastornos autoinmunes. Aquí, el sistema inmune no distingue adecuadamente entre uno mismo y el no yo, y ataca parte del cuerpo. En circunstancias normales, muchas células T y anticuerpos reaccionan con péptidos "propios". Una de las funciones de las células especializadas (ubicadas en el timo y la médula ósea) es presentar linfocitos jóvenes con autoantígenos producidos en todo el cuerpo y eliminar las células que reconocen autoantígenos, previniendo la autoinmunidad.
Hipersensibilidad
La hipersensibilidad es una respuesta inmune que daña los propios tejidos del cuerpo. Se dividen en cuatro clases (Tipo I - IV) en función de los mecanismos implicados y el transcurso del tiempo de la reacción de hipersensibilidad. La hipersensibilidad de tipo I es una reacción inmediata o anafiláctica, a menudo asociada a la alergia. Los síntomas pueden variar desde una leve molestia hasta la muerte. La hipersensibilidad de tipo I está mediada por IgE, que desencadena la desgranulación de mastocitos y basófilos cuando se unen por antígeno. La hipersensibilidad tipo II ocurre cuando los anticuerpos se unen a los antígenos en las propias células del paciente, marcándolos para su destrucción. Esto también se llama hipersensibilidad dependiente de anticuerpos (o citotóxica) y está mediada por anticuerpos IgG e IgM. Complejos inmunes (agregaciones de antígenos, proteínas del complemento, y anticuerpos IgG e IgM) depositados en diversos tejidos desencadenan reacciones de hipersensibilidad de Tipo III. Hipersensibilidad tipo IV (también conocida como mediada por células o hipersensibilidad de tipo retardado ) por lo general toma entre dos y tres días para desarrollarse. Las reacciones de tipo IV están involucradas en muchas enfermedades autoinmunes e infecciosas, pero también pueden involucrar dermatitis de contacto (hiedra venenosa). Estas reacciones están mediadas por células T, monocitos y macrófagos.
Inflamación idiopática
La inflamación es una de las primeras respuestas del sistema inmune a la infección, pero puede aparecer sin causa conocida.
La inflamación es producida por eicosanoides y citocinas, que son liberadas por las células lesionadas o infectadas. Los eicosanoides incluyen las prostaglandinas que producen fiebre y la dilatación de los vasos sanguíneos asociada a la inflamación, y los leucotrienos que atraen a ciertos glóbulos blancos (leucocitos). Las citocinas comunes incluyen interleuquinas que son responsables de la comunicación entre los glóbulos blancos; quimiocinas que promueven la quimiotaxis; e interferones que tienen efectos antivirales, como el cierre de la síntesis de proteínas en la célula huésped. Los factores de crecimiento y los factores citotóxicos también pueden ser liberados. Estas citoquinas y otras sustancias químicas reclutan células inmunitarias en el sitio de infección y promueven la cicatrización de cualquier tejido dañado después de la eliminación de patógenos.
Otros mecanismos y evolución
Es probable que surgiera un sistema inmune adaptativo multicomponente con los primeros vertebrados, ya que los invertebrados no generan linfocitos o una respuesta humoral basada en anticuerpos. Muchas especies, sin embargo, utilizan mecanismos que parecen ser precursores de estos aspectos de la inmunidad de vertebrados. Los sistemas inmunitarios aparecen incluso en las formas de vida estructuralmente más simples, con bacterias que usan un mecanismo de defensa único, llamado sistema de modificación de restricción para protegerse de los patógenos virales, llamados bacteriófagos. Los procariotas también poseen inmunidad adquirida, a través de un sistema que usa secuencias CRISPR para retener fragmentos de los genomas de los fagos con los que han entrado en contacto en el pasado, lo que les permite bloquear la replicación del virus a través de una forma de interferencia de ARN. Los procariotas también poseen otros mecanismos de defensa.
Los receptores de reconocimiento de patrón son proteínas utilizadas por casi todos los organismos para identificar moléculas asociadas con patógenos. Los péptidos antimicrobianos llamados defensinas son un componente evolutivamente conservado de la respuesta inmune innata que se encuentra en todos los animales y plantas, y representan la principal forma de inmunidad sistémica de los invertebrados. El sistema del complemento y las células fagocíticas también se utilizan en la mayoría de las formas de vida de los invertebrados. Las ribonucleasas y la vía de interferencia de ARN se conservan en todos los eucariotas, y se cree que juegan un papel en la respuesta inmune a los virus.
A diferencia de los animales, las plantas carecen de células fagocíticas, pero muchas respuestas inmunitarias de las plantas implican señales químicas sistémicas que se envían a través de una planta. Las células vegetales individuales responden a moléculas asociadas con patógenos conocidos como patrones moleculares asociados a patógenos o PAMP. Cuando una parte de una planta se infecta, la planta produce una respuesta hipersensible localizada, mediante la cual las células en el sitio de la infección experimentan apoptosis rápida para evitar la propagación de la enfermedad a otras partes de la planta. La resistencia sistémica adquirida (SAR) es un tipo de respuesta defensiva utilizada por las plantas que hace que toda la planta sea resistente a un agente infeccioso en particular. Los mecanismos de silenciamiento de ARN son particularmente importantes en esta respuesta sistémica, ya que pueden bloquear la replicación del virus.
Inmunología tumoral

Otro papel importante del sistema inmune es identificar y eliminar tumores. Esto se llama vigilancia inmune . Las células transformadas de tumores expresan antígenos que no se encuentran en las células normales. Para el sistema inmune, estos antígenos parecen extraños, y su presencia hace que las células inmunes ataquen las células tumorales transformadas. Los antígenos expresados por tumores tienen varias fuentes; algunos se derivan de virus oncogénicos como el virus del papiloma humano, que causa cáncer de cuello uterino, mientras que otros son proteínas propias del organismo que se producen en niveles bajos en células normales pero alcanzan niveles elevados en células tumorales. Un ejemplo es una enzima llamada tirosinasa que, cuando se expresa en niveles altos, transforma ciertas células de la piel (por ejemplo, melanocitos) en tumores llamados melanomas. Una tercera fuente posible de antígenos tumorales son proteínas normalmente importantes para regular el crecimiento y la supervivencia celular, que comúnmente mutan en moléculas inductoras de cáncer llamadas oncogenes.
La principal respuesta del sistema inmune a los tumores es destruir las células anormales utilizando células T asesinas, a veces con la ayuda de las células T auxiliares. Los antígenos tumorales se presentan en moléculas MHC de clase I de forma similar a los antígenos virales. Esto permite que las células T asesinas reconozcan la célula tumoral como anormal. Las células NK también matan las células tumorales de una manera similar, especialmente si las células tumorales tienen menos moléculas MHC de clase I en su superficie de lo normal; este es un fenómeno común con los tumores. En ocasiones, se generan anticuerpos contra células tumorales que permiten su destrucción por el sistema del complemento.
Claramente, algunos tumores evaden el sistema inmune y se convierten en cáncer. Las células tumorales a menudo tienen un número reducido de moléculas MHC de clase I en su superficie, evitando así la detección por las células T asesinas. Algunas células tumorales también liberan productos que inhiben la respuesta inmune; por ejemplo, secretando la citocina TGF-β, que suprime la actividad de macrófagos y linfocitos. Además, la tolerancia inmunológica puede desarrollarse contra los antígenos tumorales, por lo que el sistema inmunitario ya no ataca las células tumorales.
Paradójicamente, los macrófagos pueden promover el crecimiento tumoral cuando las células tumorales envían citoquinas que atraen a los macrófagos, que luego generan citoquinas y factores de crecimiento como el factor alfa de necrosis tumoral que nutren el desarrollo tumoral o promueven la plasticidad de células madre. Además, una combinación de hipoxia en el tumor y una citoquina producida por los macrófagos induce a las células tumorales a disminuir la producción de una proteína que bloquea la metástasis y ayuda a la diseminación de las células cancerosas.
Regulación fisiológica
El sistema inmune está involucrado en muchos aspectos de la regulación fisiológica en el cuerpo. El sistema inmune interactúa íntimamente con otros sistemas, como el sistema endocrino y el nervioso. El sistema inmune también juega un papel crucial en la embriogénesis (desarrollo del embrión), así como en la reparación y regeneración de tejidos.
Hormonas
Las hormonas pueden actuar como inmunomoduladores, alterando la sensibilidad del sistema inmune. Por ejemplo, las hormonas sexuales femeninas son inmunoestimuladores conocidos tanto de respuestas inmunitarias adaptadas como innatas. Algunas enfermedades autoinmunes como el lupus eritematoso afectan a las mujeres de manera preferencial, y su aparición a menudo coincide con la pubertad. Por el contrario, las hormonas sexuales masculinas, como la testosterona, parecen ser inmunosupresoras. Otras hormonas parecen regular también el sistema inmunitario, en particular la prolactina, la hormona de crecimiento y la vitamina D.
Vitamina D
Cuando una célula T encuentra un patógeno extraño, extiende un receptor de vitamina D. Esto es esencialmente un dispositivo de señalización que permite que las células T se unan a la forma activa de la vitamina D, la hormona esteroidea calcitriol. Las células T tienen una relación simbiótica con la vitamina D. Las células T no solo extienden un receptor de vitamina D, en esencia, solicitan unirse a la versión de la hormona esteroidea de la vitamina D, calcitriol, sino que las células T expresan el gen CYP27B1. que es el gen responsable de convertir la versión prehormonal de la vitamina D, calcidiol en la versión de la hormona esteroidea, calcitriol. Solo después de unirse al calcitriol, las células T pueden realizar su función prevista. Otras células del sistema inmune que se sabe que expresan CYP27B1 y por lo tanto activan el calcidiol de vitamina D, son células dendríticas, queratinocitos y macrófagos.
Se conjetura que una disminución progresiva en los niveles de la hormona con la edad es parcialmente responsable de la respuesta inmune debilitada en los individuos que envejecen. Por el contrario, algunas hormonas están reguladas por el sistema inmune, especialmente la actividad de la hormona tiroidea. El declive relacionado con la edad en la función inmune también está relacionado con la disminución de los niveles de vitamina D en los ancianos. A medida que las personas envejecen, suceden dos cosas que afectan negativamente sus niveles de vitamina D. En primer lugar, permanecen en el interior más debido a la disminución de los niveles de actividad. Esto significa que obtienen menos sol y, por lo tanto, producen menos colecalciferol a través de la radiación UVB. En segundo lugar, a medida que una persona envejece, la piel se vuelve menos experta en producir vitamina D.
Duerme y descansa
El sistema inmune se ve afectado por el sueño y el descanso, y la privación del sueño es perjudicial para la función inmune. Los complejos circuitos de retroalimentación que involucran a las citocinas, como la interleucina-1 y el factor de necrosis tumoral α producidos en respuesta a la infección, también parecen desempeñar un papel en la regulación del sueño de movimiento ocular no rápido (REM). Por lo tanto, la respuesta inmune a la infección puede provocar cambios en el ciclo del sueño, incluido un aumento en el sueño de onda lenta en relación con el sueño REM.
Cuando sufren de falta de sueño, las inmunizaciones activas pueden tener un efecto disminuido y pueden dar como resultado una producción de anticuerpos más baja y una respuesta inmune más baja que la que se observaría en un individuo bien descansado. Además, proteínas como NFIL3, que se ha demostrado que están estrechamente entrelazadas con la diferenciación de células T y nuestros ritmos circadianos, pueden verse afectadas por la alteración de la luz natural y los ciclos oscuros a través de la privación de sueño, el trabajo por turnos, etc. como resultado, estas interrupciones pueden conducir a un aumento de las enfermedades crónicas, como las enfermedades del corazón, el dolor crónico y el asma.
Además de las consecuencias negativas de la privación del sueño, el sueño y el sistema circadiano entrelazado han demostrado tener fuertes efectos reguladores sobre las funciones inmunológicas que afectan tanto a la inmunidad innata como a la adaptativa. En primer lugar, durante la etapa temprana de sueño de onda lenta, una caída repentina en los niveles sanguíneos de cortisol, epinefrina y norepinefrina inducen un aumento de los niveles sanguíneos de las hormonas leptina, hipofisaria y prolactina. Estas señales inducen un estado proinflamatorio a través de la producción de las citocinas proinflamatorias interleucina-1, interleucina-12, TNF-alfa e IFN-gamma. Estas citoquinas luego estimulan las funciones inmunitarias, como la activación, proliferación y diferenciación de las células inmunes. Es durante este tiempo que las células T de memoria indiferenciadas, o menos diferenciadas, como ingenuas y centrales, pico (es decir, durante un tiempo de una respuesta inmune adaptativa que evoluciona lentamente). Además de estos efectos, el entorno de las hormonas producidas en este momento (leptina, hormona de crecimiento hipofisaria y prolactina) respalda las interacciones entre APC y células T, un cambio de la T
h 1 / T
h 2 equilibrio de citoquinas hacia uno que soporta T h 1, un aumento en la proliferación global de células T h , y la migración de linfocitos T a nódulos linfáticos. También se cree que este medio ayuda a la formación de memoria inmune duradera a través del inicio de las respuestas inmunitarias Th1.
h 1 / T
h 2 equilibrio de citoquinas hacia uno que soporta T h 1, un aumento en la proliferación global de células T h , y la migración de linfocitos T a nódulos linfáticos. También se cree que este medio ayuda a la formación de memoria inmune duradera a través del inicio de las respuestas inmunitarias Th1.
Por el contrario, durante los períodos de vigilia las células efectoras diferenciadas, tales como las células asesinas naturales citotóxicas y los CTL (linfocitos T citotóxicos), alcanzan el máximo para provocar una respuesta eficaz contra cualquier patógeno intrusivo. Además durante los tiempos activos de despertar, las moléculas antiinflamatorias, como el cortisol y las catecolaminas, alcanzan un pico. Hay dos teorías sobre por qué el estado proinflamatorio está reservado para el tiempo de sueño. En primer lugar, la inflamación causaría deficiencias cognitivas y físicas graves si ocurriera durante los períodos de vigilia. En segundo lugar, la inflamación puede ocurrir durante los períodos de sueño debido a la presencia de melatonina. La inflamación causa una gran cantidad de estrés oxidativo y la presencia de melatonina durante los períodos de sueño podría contrarrestar activamente la producción de radicales libres durante este tiempo.
Nutrición y dieta
La desnutrición se asocia con enfermedades como la diabetes y la obesidad, que se sabe que afectan la función inmune. Una desnutrición más moderada, así como ciertas deficiencias específicas de oligoelementos y nutrientes, también pueden comprometer la respuesta inmune.
Los alimentos ricos en ciertos ácidos grasos pueden fomentar un sistema inmunológico saludable. Del mismo modo, la desnutrición fetal puede causar una discapacidad permanente del sistema inmunitario.
Reparación y regeneración
El sistema inmune, particularmente el componente innato, juega un papel decisivo en la reparación de los tejidos después de un insulto. Los actores clave incluyen macrófagos y neutrófilos, pero también son importantes otros actores celulares, como las células T γδ, las células linfoides innatas (ILC) y las células T reguladoras (Tregs). La plasticidad de las células inmunitarias y el equilibrio entre las señales proinflamatorias y antiinflamatorias son aspectos cruciales de la reparación eficiente de los tejidos. Los componentes y vías inmunes también están involucrados en la regeneración, por ejemplo en anfibios. Según una hipótesis, los organismos que pueden regenerarse podrían ser menos inmunocompetentes que los organismos que no pueden regenerarse.
Manipulación en medicina
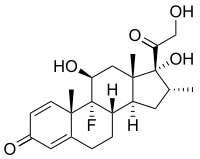
La respuesta inmune puede ser manipulada para suprimir las respuestas no deseadas que resultan de la autoinmunidad, la alergia y el rechazo de trasplantes, y para estimular respuestas protectoras contra patógenos que en gran medida eluden el sistema inmune (ver inmunización) o el cáncer.
Inmunosupresión
Los medicamentos inmunosupresores se usan para controlar los trastornos autoinmunes o la inflamación cuando se produce un daño excesivo en el tejido y para evitar el rechazo del trasplante después de un trasplante de órgano.
Los medicamentos antiinflamatorios a menudo se utilizan para controlar los efectos de la inflamación. Los glucocorticoides son los más poderosos de estos medicamentos; sin embargo, estos medicamentos pueden tener muchos efectos secundarios indeseables, como obesidad central, hiperglucemia, osteoporosis, y su uso debe controlarse estrictamente. Las dosis más bajas de medicamentos antiinflamatorios a menudo se usan junto con medicamentos citotóxicos o inmunosupresores como el metotrexato o la azatioprina. Los fármacos citotóxicos inhiben la respuesta inmune al matar las células en división, como las células T activadas. Sin embargo, la matanza es indiscriminada y otras células que se dividen constantemente y sus órganos se ven afectados, lo que causa efectos secundarios tóxicos. Los fármacos inmunosupresores, como la ciclosporina, evitan que las células T respondan correctamente a las señales al inhibir las vías de transducción de señales.
Inmunoestimulación
La inmunoterapia contra el cáncer cubre las formas médicas de estimular el sistema inmune para atacar los tumores cancerosos.
Acercamientos teóricos al sistema inmune
La inmunología es muy experimental en la práctica diaria, pero también se caracteriza por una actitud teórica constante. Se han sugerido muchas teorías en inmunología desde fines del siglo XIX hasta la actualidad. El final del siglo 19 y el comienzo del siglo 20 vio una batalla entre las teorías de la inmunidad "celular" y "humoral". De acuerdo con la teoría celular de la inmunidad, representada en particular por Elie Metchnikoff, fueron las células, más precisamente, los fagocitos, las responsables de la respuesta inmune. Por el contrario, la teoría humoral de la inmunidad, sostenida, entre otros, por Robert Koch y Emil von Behring, declaró que los agentes inmunes activos eran componentes solubles (moléculas) que se encuentran en los "humores" del organismo en lugar de sus células.
A mediados de la década de 1950, Frank Burnet, inspirado en una sugerencia hecha por Niels Jerne, formuló la teoría de la selección clonal (CST) de la inmunidad. Sobre la base del CST, Burnet desarrolló una teoría de cómo se desencadena una respuesta inmune según la distinción entre uno mismo y el otro: los constituyentes "propios" (constituyentes del cuerpo) no desencadenan respuestas inmunes destructivas, mientras que las entidades "no propias" (patógenos, un aloinjerto) desencadena una respuesta inmune destructiva. La teoría se modificó más tarde para reflejar nuevos descubrimientos con respecto a la histocompatibilidad o la activación compleja de "dos señales" de las células T. Se ha criticado la teoría de la inmunidad y el vocabulario de sí mismo / no solo del yo / no yo mismo, pero sigue siendo muy influyente.
Más recientemente, se han sugerido varios marcos teóricos en inmunología, que incluyen puntos de vista "autopoiéticos", puntos de vista "inmunes cognitivos", el "modelo de peligro" (o "teoría del peligro") y la teoría de la "discontinuidad". El modelo de peligro, sugerido por Polly Matzinger y sus colegas, ha sido muy influyente, despertando muchos comentarios y discusiones.
Predecir la inmunogenicidad
Los medicamentos más grandes (> 500 Da) pueden provocar una respuesta inmune neutralizante, especialmente si los medicamentos se administran repetidamente o en dosis mayores. Esto limita la efectividad de los medicamentos basados en péptidos y proteínas más grandes (que típicamente son más grandes que 6000 Da). En algunos casos, el medicamento en sí no es inmunogénico, pero puede administrarse conjuntamente con un compuesto inmunogénico, como a veces es el caso del Taxol. Se han desarrollado métodos computacionales para predecir la inmunogenicidad de péptidos y proteínas, que son particularmente útiles en el diseño de anticuerpos terapéuticos, la evaluación de la posible virulencia de mutaciones en partículas de recubrimiento viral y la validación de tratamientos de fármacos basados en péptidos propuestos. Las primeras técnicas se basaban principalmente en la observación de que los aminoácidos hidrofílicos están sobrerrepresentados en regiones epitópicas que los aminoácidos hidrofóbicos; sin embargo, los desarrollos más recientes se basan en técnicas de aprendizaje automático que utilizan bases de datos de epítopos conocidos existentes, generalmente sobre proteínas de virus bien estudiadas, como un conjunto de entrenamiento. Se ha establecido una base de datos de acceso público para la catalogación de epítopos de patógenos que se sabe son reconocibles por las células B. El campo emergente de los estudios de inmunogenicidad basados en la bioinformática se conoce como inmunoinformática . La inmunoproteómica es el estudio de grandes conjuntos de proteínas (proteómica) involucradas en la respuesta inmune.
Manipulación por patógenos
El éxito de cualquier patógeno depende de su capacidad para eludir las respuestas inmunitarias del huésped. Por lo tanto, los patógenos desarrollaron varios métodos que les permiten infectar con éxito a un huésped, al tiempo que evitan la detección o destrucción por parte del sistema inmune. Las bacterias a menudo superan barreras físicas secretando enzimas que digieren la barrera, por ejemplo, usando un sistema de secreción tipo II. Alternativamente, usando un sistema de secreción tipo III, pueden insertar un tubo hueco en la célula huésped, proporcionando una ruta directa para que las proteínas se muevan del patógeno al huésped. Estas proteínas a menudo se utilizan para cerrar las defensas del huésped.
Una estrategia de evasión utilizada por varios patógenos para evitar el sistema inmune innato es ocultarse dentro de las células de su huésped (también llamada patogénesis intracelular). Aquí, un patógeno pasa la mayor parte de su ciclo de vida dentro de las células del huésped, donde está protegido del contacto directo con las células inmunes, los anticuerpos y el complemento. Algunos ejemplos de patógenos intracelulares incluyen virus, la bacteria de intoxicación alimentaria Salmonella y los parásitos eucarióticos que causan la malaria ( Plasmodium falciparum ) y la leishmaniasis ( Leishmania spp. ). Otras bacterias, como Mycobacterium tuberculosis, vive dentro de una cápsula protectora que impide la lisis por complemento. Muchos patógenos secretan compuestos que disminuyen o dirigen mal la respuesta inmune del huésped. Algunas bacterias forman biofilms para protegerse de las células y las proteínas del sistema inmune. Tales biofilms están presentes en muchas infecciones exitosas, por ejemplo, las infecciones crónicas de Pseudomonas aeruginosa y Burkholderia cenocepacia características de la fibrosis quística. Otras bacterias generan proteínas de superficie que se unen a los anticuerpos y los vuelven ineficaces; Los ejemplos incluyen Streptococcus (proteína G), Staphylococcus aureus (proteína A) y Peptostreptococcus magnus (proteína L).
Los mecanismos utilizados para evadir el sistema inmune adaptativo son más complicados. El enfoque más simple es cambiar rápidamente epítopos no esenciales (aminoácidos y / o azúcares) en la superficie del patógeno, mientras se mantienen ocultos los epítopos esenciales. Esto se llama variación antigénica. Un ejemplo es el VIH, que muta rápidamente, por lo que las proteínas en su envoltura viral que son esenciales para la entrada en su célula diana huésped están cambiando constantemente. Estos frecuentes cambios en los antígenos pueden explicar los fallos de las vacunas dirigidas a este virus. El parásito Trypanosoma brucei utiliza una estrategia similar, cambiando constantemente un tipo de proteína de superficie por otra, lo que le permite mantenerse un paso por delante de la respuesta de anticuerpos. Enmascarar antígenos con moléculas hospedadoras es otra estrategia común para evitar la detección por el sistema inmune. En el VIH, la envoltura que cubre el virión se forma a partir de la membrana más externa de la célula huésped; tales virus "autoencubiertos" dificultan que el sistema inmune los identifique como estructuras "no propias".