Virus
Definición
Virus | |
|---|---|
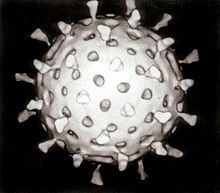 | |
| Rotavirus | |
Desde el artículo de 1893 de Dmitri Ivanovsky que describe un patógeno no bacteriano que infecta plantas de tabaco y el descubrimiento del virus del mosaico del tabaco por Martinus Beijerinck en 1898, se han descrito en detalle alrededor de 5.000 especies de virus, aunque hay millones de tipos. Los virus se encuentran en casi todos los ecosistemas de la Tierra y son el tipo más grande de entidad biológica. El estudio de los virus se conoce como virología, una subespecialidad de la microbiología.
Mientras no estén dentro de una célula infectada o en el proceso de infectar una célula, los virus existen en forma de partículas independientes. Estas partículas virales , también conocidas como viriones , consisten en: (i) el material genético hecho a partir de ADN o ARN, moléculas largas que llevan información genética; (ii) un impermeable, llamado cápside, que rodea y protege el material genético; y en algunos casos (iii) una envoltura de lípidos que rodea el recubrimiento de proteína. Las formas de estas partículas de virus van desde simples formas helicoidales e icosaédricas para algunas especies de virus hasta estructuras más complejas para otras. La mayoría de las especies de virus tienen viriones que son demasiado pequeños para ser vistos con un microscopio óptico. El virión promedio es aproximadamente una centésima del tamaño de una bacteria promedio.
Los orígenes de los virus en la historia evolutiva de la vida no están claros: algunos pueden haber evolucionado a partir de plásmidos, pedazos de ADN que pueden moverse entre las células, mientras que otros pueden haber evolucionado a partir de las bacterias. En la evolución, los virus son un medio importante de transferencia horizontal de genes, que aumenta la diversidad genética. Los virus son considerados por algunos como una forma de vida, ya que transportan material genético, se reproducen y evolucionan a través de la selección natural, pero carecen de características clave (como la estructura celular) que generalmente se consideran necesarias para contar como vida. Debido a que poseen algunas de estas cualidades, pero no todas, los virus se han descrito como "organismos al borde de la vida" y como replicadores.
Los virus se diseminan de muchas maneras; los virus en las plantas a menudo se transmiten de una planta a otra por insectos que se alimentan de la savia de la planta, como los pulgones; los virus en animales pueden ser transportados por insectos chupadores de sangre. Estos organismos portadores de enfermedades son conocidos como vectores. Los virus de la influenza se transmiten al toser y estornudar. El norovirus y el rotavirus, causas comunes de gastroenteritis viral, se transmiten por vía fecal-oral y se transmiten de persona a persona por contacto, ingresando al organismo en alimentos o agua. El VIH es uno de varios virus transmitidos por contacto sexual y por exposición a sangre infectada. La variedad de células anfitrionas que un virus puede infectar se llama su "rango de host". Esto puede ser limitado, lo que significa que un virus es capaz de infectar a pocas especies, o amplio, lo que significa que es capaz de infectar a muchos.
Las infecciones virales en animales provocan una respuesta inmune que generalmente elimina el virus infectante. Las respuestas inmunes también pueden producirse mediante vacunas, que confieren una inmunidad adquirida artificialmente a la infección viral específica. Algunos virus, incluidos los que causan el SIDA y la hepatitis viral, eluden estas respuestas inmunitarias y provocan infecciones crónicas. Los antibióticos no tienen ningún efecto sobre los virus, pero se han desarrollado varios medicamentos antivirales.
Etimología
La palabra es del latín neutro vīrus que se refiere al veneno y otros líquidos nocivos, de "la misma base indoeuropea que el veneno sánscrito viṣa , Avestan vīša veneno, griego antiguo ἰός veneno", atestiguado por primera vez en inglés en 1398 en la traducción de John Trevisa de De Proprietatibus Rerum de Bartholomeus Anglicus . Virulento , del latín virulentus (venenoso), data de c. 1400. Un significado de "agente que causa enfermedades infecciosas" se registra por primera vez en 1728, antes del descubrimiento de virus por Dmitri Ivanovsky en 1892. El plural en inglés es virus (a veces también viri o vira).), mientras que la palabra latina es un sustantivo masivo, que no tiene un plural clásicamente atestiguado ( vīra se usa en neolatino). El adjetivo viral data de 1948. El término virion ( viriones plural ), que data de 1959, también se usa para referirse a una sola partícula viral infecciosa estable que se libera de la célula y es totalmente capaz de infectar otras células de la misma tipo.
Historia

Louis Pasteur no pudo encontrar un agente causante de la rabia y especuló sobre un patógeno demasiado pequeño para detectarlo con un microscopio. En 1884, el microbiólogo francés Charles Chamberland inventó un filtro (conocido hoy como el filtro Chamberland o el filtro Pasteur-Chamberland) con poros más pequeños que las bacterias. Por lo tanto, podría pasar una solución que contiene bacterias a través del filtro y eliminarlas por completo. En 1892, el biólogo ruso Dmitri Ivanovsky usó este filtro para estudiar lo que ahora se conoce como el virus del mosaico del tabaco. Sus experimentos demostraron que los extractos de hojas trituradas de plantas de tabaco infectadas permanecen infecciosas después de la filtración. Ivanovsky sugirió que la infección podría ser causada por una toxina producida por bacterias, pero no persiguió la idea. En ese momento se pensó que todos los agentes infecciosos podían retenerse mediante filtros y cultivarse en un medio nutriente; esto era parte de la teoría de los gérmenes de la enfermedad. En 1898, el microbiólogo holandés Martinus Beijerinck repitió los experimentos y se convenció de que la solución filtrada contenía una nueva forma de agente infeccioso. Observó que el agente se multiplicaba solo en las células que se estaban dividiendo, pero como sus experimentos no demostraban que estuviera hecho de partículas, lo llamó contagium vivum fluidum (germen viviente soluble) y reintrodujo la palabra virus . Beijerinck sostuvo que los virus eran de naturaleza líquida, una teoría desacreditada posteriormente por Wendell Stanley, quien demostró que eran partículas. En el mismo año, Friedrich Loeffler y Paul Frosch pasaron el primer virus animal - agente de la fiebre aftosa (aphthovirus) - a través de un filtro similar.
A principios del siglo XX, el bacteriólogo inglés Frederick Twort descubrió un grupo de virus que infectan bacterias, ahora llamadas bacteriófagos (o comúnmente fagos).), y el microbiólogo franco-canadiense Félix d'Herelle describió virus que, cuando se agregan a bacterias en una placa de agar, producirían áreas de bacterias muertas. Él diluyó con precisión una suspensión de estos virus y descubrió que las diluciones más altas (concentraciones de virus más bajas), en lugar de matar a todas las bacterias, formaban áreas discretas de organismos muertos. Contar estas áreas y multiplicar por el factor de dilución le permitió calcular la cantidad de virus en la suspensión original. Los fagos fueron anunciados como un tratamiento potencial para enfermedades como la tifoidea y el cólera, pero su promesa fue olvidada con el desarrollo de la penicilina. El estudio de fagos proporcionó información sobre la activación y desactivación de genes y un mecanismo útil para introducir genes extraños en las bacterias.
A finales del siglo XIX, los virus se definieron en términos de su infectividad, su capacidad de ser filtrados y su requisito de hospedadores vivos. Los virus se habían cultivado solo en plantas y animales. En 1906, Ross Granville Harrison inventó un método para cultivar tejido en la linfa y, en 1913, E. Steinhardt, C. Israeli y RA Lambert utilizaron este método para cultivar el virus vaccinia en fragmentos de tejido corneal de conejillo de Indias. En 1928, HB Maitland y MC Maitland crecieron el virus vaccinia en suspensiones de riñones de gallinas picadas. Su método no fue ampliamente adoptado hasta la década de 1950, cuando el poliovirus se cultivó a gran escala para la producción de vacunas.
Otro avance se produjo en 1931, cuando el patólogo estadounidense Ernest William Goodpasture y Alice Miles Woodruff crecieron la gripe y varios otros virus en huevos de gallina fertilizados. En 1949, John Franklin Enders, Thomas Weller y Frederick Robbins crecieron el virus de la polio en células embrionarias humanas cultivadas, el primer virus que se cultiva sin usar tejido animal sólido o huevos. Este trabajo permitió a Jonas Salk hacer una vacuna eficaz contra la polio.
Las primeras imágenes de virus se obtuvieron con la invención del microscopio electrónico en 1931 por los ingenieros alemanes Ernst Ruska y Max Knoll. En 1935, el bioquímico y virólogo estadounidense Wendell Meredith Stanley examinó el virus del mosaico del tabaco y descubrió que en su mayoría estaba hecho de proteínas. Poco tiempo después, este virus se separó en partes de proteína y ARN. El virus del mosaico del tabaco fue el primero en cristalizarse y, por lo tanto, su estructura pudo elucidarse en detalle. Bernal y Fankuchen obtuvieron las primeras imágenes de difracción de rayos X del virus cristalizado en 1941. Sobre la base de sus imágenes, Rosalind Franklin descubrió la estructura completa del virus en 1955. En el mismo año,
La segunda mitad del siglo 20 fue la edad de oro del descubrimiento de virus y la mayoría de las más de 2.000 especies reconocidas de animales, plantas y virus bacterianos fueron descubiertos durante estos años. En 1957, se descubrió el arterivirus equino y la causa de la diarrea vírica bovina (un pestivirus). En 1963, el virus de la hepatitis B fue descubierto por Baruch Blumberg, y en 1965, Howard Temin describió el primer retrovirus. La transcriptasa inversa, la enzima que utilizan los retrovirus para hacer copias de ADN de su ARN, fue descrita por primera vez en 1970, independientemente por Howard Martin Temin y David Baltimore. En 1983, el equipo de Luc Montagnier en el Instituto Pasteur en Francia, primero aisló el retrovirus ahora llamado VIH. En 1989, el equipo de Michael Houghton en Chiron Corporation descubrió Hepatitis C.
Orígenes
Los virus se encuentran dondequiera que haya vida y probablemente hayan existido desde que las células vivas evolucionaron por primera vez. El origen de los virus no está claro porque no forman fósiles, por lo que las técnicas moleculares se han utilizado para comparar el ADN o ARN de los virus y son un medio útil para investigar cómo surgieron. Además, el material genético viral puede ocasionalmente integrarse en la línea germinal de los organismos hospedadores, por lo que pueden transmitirse verticalmente a la descendencia del huésped durante muchas generaciones. Esto proporciona una fuente inestimable de información para que los paleovirus rastreen antiguos virus que existieron hace millones de años. Hay tres hipótesis principales que pretenden explicar los orígenes de los virus:
- Hipótesis regresiva
- Los virus pueden haber sido alguna vez células pequeñas que parasitaron células más grandes. Con el tiempo, los genes no requeridos por su parasitismo se perdieron. Las bacterias rickettsia y clamidia son células vivas que, como los virus, pueden reproducirse solo dentro de las células del huésped. Apoyan esta hipótesis, ya que su dependencia del parasitismo es probable que haya causado la pérdida de genes que les permitió sobrevivir fuera de una célula. Esto también se llama hipótesis de degeneración o hipótesis de reducción .
- Hipótesis de origen celular
- Algunos virus pueden haber evolucionado a partir de fragmentos de ADN o ARN que "escaparon" de los genes de un organismo más grande. El ADN escapado podría provenir de plásmidos (trozos de ADN desnudo que pueden moverse entre las células) o transposones (moléculas de ADN que se replican y se mueven a diferentes posiciones dentro de los genes de la célula). Una vez denominados "genes saltarines", los transposones son ejemplos de elementos genéticos móviles y podrían ser el origen de algunos virus. Fueron descubiertos en el maíz por Barbara McClintock en 1950. Esto a veces se llama la hipótesis de la vagancia , o la hipótesis de escape .
- Hipótesis de co-evolución
- Esto también se llama la hipótesis del virus primero y propone que los virus pueden haber evolucionado a partir de moléculas complejas de proteína y ácido nucleico al mismo tiempo que las células aparecieron por primera vez en la Tierra y habrían dependido de la vida celular durante miles de millones de años. Los viroides son moléculas de ARN que no están clasificadas como virus porque carecen de una cubierta de proteína. Tienen características que son comunes a varios virus y a menudo se llaman agentes subvirales. Los viroides son importantes patógenos de las plantas. No codifican proteínas, sino que interactúan con la célula anfitriona y utilizan la maquinaria de host para su replicación. El virus de la hepatitis delta de los seres humanos tiene un genoma de ARN similar a los viroides, pero tiene una cubierta proteica derivada del virus de la hepatitis B y no puede producir uno propio. Es, por lo tanto, un virus defectuoso. Aunque el genoma del virus de la hepatitis delta puede replicarse independientemente una vez dentro de una célula huésped, requiere la ayuda del virus de la hepatitis B para proporcionar una capa de proteína para que pueda transmitirse a nuevas células. De manera similar, el virophage sputnik depende del mimivirus, que infecta el protozoo Acanthamoeba castellanii . Estos virus, que dependen de la presencia de otras especies de virus en la célula huésped, se llaman satélites y pueden representar intermedios evolutivos de viroides y virus.
En el pasado, había problemas con todas estas hipótesis: la hipótesis regresiva no explicaba por qué incluso los parásitos celulares más pequeños no se asemejan a los virus de ninguna manera. La hipótesis de escape no explica las cápsides complejas y otras estructuras en partículas de virus. La hipótesis del virus contravino la definición de virus en la medida en que requieren células hospedadoras. Los virus ahora son reconocidos como antiguos y tienen orígenes anteriores a la divergencia de vida en los tres dominios. Este descubrimiento ha llevado a los virólogos modernos a reconsiderar y reevaluar estas tres hipótesis clásicas.
La evidencia de un mundo ancestral de células de ARN y el análisis por computadora de las secuencias de ADN víricas y de hospedador están dando una mejor comprensión de las relaciones evolutivas entre diferentes virus y pueden ayudar a identificar a los antepasados de los virus modernos. Hasta la fecha, tales análisis no han probado cuál de estas hipótesis es correcta. Parece poco probable que todos los virus actualmente conocidos tengan un ancestro común, y los virus probablemente hayan surgido numerosas veces en el pasado por uno o más mecanismos.
Los priones son moléculas de proteínas infecciosas que no contienen ADN ni ARN. Pueden causar infecciones como la tembladera en las ovejas, la encefalopatía espongiforme bovina (enfermedad de las "vacas locas") en el ganado y la enfermedad de desgaste crónica en los ciervos; en humanos, las enfermedades priónicas incluyen Kuru, enfermedad de Creutzfeldt-Jakob y síndrome de Gerstmann-Sträussler-Scheinker. Aunque los priones son fundamentalmente diferentes de los virus y viroides, su descubrimiento da credibilidad a la teoría de que los virus podrían haber evolucionado a partir de moléculas autorreplicantes.
Microbiología
Propiedades de la vida
Las opiniones difieren sobre si los virus son una forma de vida o estructuras orgánicas que interactúan con organismos vivos. Se han descrito como "organismos al borde de la vida", ya que se asemejan a los organismos en el sentido de que poseen genes, evolucionan por selección natural y se reproducen creando copias múltiples de sí mismos mediante el autoensamblaje. Aunque tienen genes, no tienen una estructura celular, que a menudo se considera la unidad básica de la vida. Los virus no tienen su propio metabolismo y requieren una célula huésped para fabricar nuevos productos. Por lo tanto, no pueden reproducirse naturalmente fuera de una célula huésped, aunque especies bacterianas como la rickettsia y la clamidia se consideran organismos vivos a pesar de la misma limitación. Las formas de vida aceptadas usan la división celular para reproducirse, mientras que los virus se ensamblan espontáneamente dentro de las células. Difieren del crecimiento autónomo de los cristales ya que heredan mutaciones genéticas mientras están sujetos a la selección natural. El autoensamblaje del virus dentro de las células anfitrionas tiene implicaciones para el estudio del origen de la vida, ya que da mayor credibilidad a la hipótesis de que la vida podría haber comenzado como moléculas orgánicas autoensambladas.
Estructura



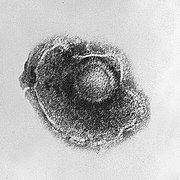
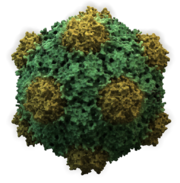
Los virus muestran una gran diversidad de formas y tamaños, llamados morfologías . En general, los virus son mucho más pequeños que las bacterias. La mayoría de los virus que se han estudiado tienen un diámetro entre 20 y 300 nanómetros. Algunos filovirus tienen una longitud total de hasta 1400 nm; sus diámetros son de solo 80 nm. La mayoría de los virus no se pueden ver con un microscopio óptico, por lo que se utilizan microscopios electrónicos de barrido y transmisión para visualizarlos. Para aumentar el contraste entre los virus y el fondo, se utilizan "tintes" densos en electrones. Estas son soluciones de sales de metales pesados, como el tungsteno, que dispersan los electrones de las regiones cubiertas por la mancha. Cuando los viriones están recubiertos con tinción (tinción positiva), los detalles finos se oscurecen. La tinción negativa supera este problema al teñir el fondo solamente.
Una partícula de virus completa, conocida como virión, consiste en ácido nucleico rodeado por una capa protectora de proteína llamada cápside. Estos se forman a partir de subunidades de proteínas idénticas llamadas capsómeros. Los virus pueden tener una "envoltura" de lípidos derivada de la membrana de la célula huésped. La cápside está hecha de proteínas codificadas por el genoma viral y su forma sirve como base para la distinción morfológica. Las subunidades de proteínas codificadas viralmente se autoensamblarán para formar una cápside, que en general requiere la presencia del genoma del virus. Los virus complejos codifican las proteínas que ayudan en la construcción de su cápside. Las proteínas asociadas con el ácido nucleico se conocen como nucleoproteínas, y la asociación de las proteínas de la cápside viral con el ácido nucleico viral se denomina nucleocápside. La cápside y la estructura entera del virus pueden sondearse mecánicamente (físicamente) mediante microscopía de fuerza atómica. En general, hay cuatro tipos principales de virus morfológicos:
- Helicoidal
- Estos virus están compuestos por un único tipo de capsómero apilado alrededor de un eje central para formar una estructura helicoidal, que puede tener una cavidad central o tubo. Esta disposición da como resultado viriones en forma de barra o filamentosos: estos pueden ser cortos y muy rígidos, o largos y muy flexibles. El material genético, en general, el ARN monocatenario, pero el ADNss en algunos casos, se une a la hélice proteínica mediante interacciones entre el ácido nucleico cargado negativamente y las cargas positivas sobre la proteína. En general, la longitud de una cápside helicoidal está relacionada con la longitud del ácido nucleico contenido en ella y el diámetro depende del tamaño y la disposición de los capsómeros. El virus del mosaico del tabaco bien estudiado es un ejemplo de virus helicoidal.
- Icosahedral
- La mayoría de los virus animales son icosaédricos o casi esféricos con simetría quiral icosaédrica. Un icosaedro regular es la forma óptima de formar un caparazón cerrado a partir de subunidades idénticas. El número mínimo de capsómeras idénticas requeridas para cada cara triangular es 3, lo que da 60 para el icosaedro. Muchos virus, como el rotavirus, tienen más de 60 capsómeros y parecen esféricos, pero retienen esta simetría. Para lograr esto, los capsómeros en los ápices están rodeados por otros cinco capsómeros y se llaman pentones. Los capsómeros en las caras triangulares están rodeados por otros seis y se denominan hexones. Los hexágonos son esencialmente planos y los pentones, que forman los 12 vértices, son curvos. La misma proteína puede actuar como la subunidad de los pentámeros y los hexámeros o puede estar compuesta de diferentes proteínas.
- Prolate
- Este es un icosaedro alargado a lo largo del eje quíntuple y es una disposición común de las cabezas de los bacteriófagos. Esta estructura se compone de un cilindro con una tapa en cada extremo.
- Sobre
- Algunas especies de virus se envuelven en una forma modificada de una de las membranas celulares, ya sea la membrana externa que rodea una célula huésped infectada o membranas internas como la membrana nuclear o el retículo endoplásmico, obteniendo así una bicapa lipídica externa conocida como envoltura viral. Esta membrana está tachonada con proteínas codificadas por el genoma viral y el genoma del huésped; la membrana lipídica y los carbohidratos presentes se originan por completo del huésped. El virus de la influenza y el VIH usan esta estrategia. La mayoría de los virus envueltos dependen de la envoltura para su infectividad.
- Complejo
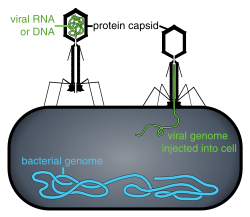
- Estos virus poseen una cápside que no es puramente helicoidal ni puramente icosaédrica, y que puede poseer estructuras extra tales como colas proteicas o una pared externa compleja. Algunos bacteriófagos, como Enterobacteria phage T4, tienen una estructura compleja que consiste en una cabeza icosaédrica unida a una cola helicoidal, que puede tener una placa base hexagonal con fibras de la cola de proteína que sobresalen. Esta estructura de cola actúa como una jeringa molecular, se une al huésped bacteriano y luego inyecta el genoma viral en la célula.
Los poxvirus son virus grandes y complejos que tienen una morfología inusual. El genoma viral se asocia con proteínas dentro de una estructura de disco central conocida como nucleoide. El nucleoide está rodeado por una membrana y dos cuerpos laterales de función desconocida. El virus tiene una envoltura externa con una gruesa capa de proteína tachonada sobre su superficie. El virión entero es levemente pleomorfo, desde ovoide a forma de ladrillo. Mimivirus es uno de los virus caracterizados más grandes, con un diámetro de cápside de 400 nm. Los filamentos de proteínas que miden 100 nm se proyectan desde la superficie. La cápside parece hexagonal bajo un microscopio electrónico, por lo tanto, la cápside es probablemente icosaédrica. En 2011, los investigadores descubrieron el virus más grande conocido en muestras de agua recolectada en el fondo del océano frente a la costa de Las Cruces, Chile. Provisionalmente llamado Megavirus chilensis , se puede ver con un microscopio óptico básico. En 2013, el género Pandoravirus se descubrió en Chile y Australia, y tiene genomas aproximadamente dos veces más grandes que Megavirus y Mimivirus.
Algunos virus que infectan Archaea tienen estructuras complejas que no están relacionadas con ninguna otra forma de virus, con una amplia variedad de formas inusuales, que van desde estructuras en forma de huso, hasta virus que se parecen a varillas enganchadas, lágrimas o incluso botellas. Otros virus de arqueas se asemejan a los bacteriófagos de cola y pueden tener múltiples estructuras de cola.
Genoma
| Propiedad | Parámetros |
|---|---|
| Ácido nucleico |
|
| Forma |
|
| Strandedness |
|
| Sentido |
|
Un virus tiene un genoma de ADN o de ARN y se denomina virus ADN o ARN, respectivamente. La gran mayoría de los virus tienen genomas de ARN. Los virus de plantas tienden a tener genomas de ARN monocatenarios y los bacteriófagos tienden a tener genomas de ADN bicatenario.
Los genomas virales son circulares , como en los poliomavirus, o lineales , como en los adenovirus. El tipo de ácido nucleico es irrelevante para la forma del genoma. Entre los virus de ARN y ciertos virus de ADN, el genoma a menudo se divide en partes separadas, en cuyo caso se denomina segmentado. Para los virus de ARN, cada segmento a menudo codifica una sola proteína y generalmente se encuentran juntos en una cápside. No se requiere que todos los segmentos estén en el mismo virión para que el virus sea infeccioso, como lo demuestra el virus mosaico brome y varios otros virus de plantas.
Un genoma viral, independientemente del tipo de ácido nucleico, casi siempre es monocatenario o bicatenario . Los genomas monocatenarios consisten en un ácido nucleico no apareado, análogo a la mitad de una escalera dividida en el medio. Los genomas bicatenarios consisten en dos ácidos nucleicos emparejados complementarios, análogos a una escalera. Las partículas de virus de algunas familias de virus, como las que pertenecen a Hepadnaviridae , contienen un genoma que es parcialmente bicatenario y parcialmente monocatenario.
Para la mayoría de los virus con genomas de ARN y algunos con genomas de ADN monocatenarios, se dice que las cadenas simples tienen sentido positivo (llamado cadena positiva ) o sentido negativo (llamada cadena negativa ), dependiendo de si son o no complementario al ARN mensajero viral (ARNm). El ARN viral de sentido positivo está en el mismo sentido que el ARNm viral y, por lo tanto, al menos una parte de él puede traducirse inmediatamente por la célula hospedadora. El ARN viral de sentido negativo es complementario al ARNm y, por lo tanto, debe convertirse en ARN de sentido positivo mediante una ARN polimerasa dependiente de ARN antes de la traducción. La nomenclatura de ADN para virus con ADN ss genómico de sentido único es similar a la nomenclatura de ARN, ya que la cadena molde para el ARNm viral es complementaria a ella (-), y el cadena de codificación es una copia de ella (+). Varios tipos de virus ssDNA y ssRNA tienen genomas que son ambisensos ya que la transcripción puede producirse en ambas cadenas en un intermediario replicativo bicatenario. Los ejemplos incluyen geminivirus, que son virus de plantas ssDNA y arenavirus, que son virus ssRNA de animales.
Tamaño del genoma
El tamaño del genoma varía mucho entre las especies. Los genomas virales más pequeños (los circovirus ADNss, familia Circoviridae ) codifican solo para dos proteínas y tienen un tamaño genómico de solo dos kilobases; los más grandes, los pandoravirus, tienen tamaños de genoma de alrededor de dos megabases que codifican unas 2500 proteínas. Los genes de los virus rara vez tienen intrones y, a menudo, están dispuestos en el genoma para que se superpongan.
En general, los virus de ARN tienen tamaños de genoma más pequeños que los virus de ADN debido a una mayor tasa de error cuando se replican, y tienen un límite máximo de tamaño máximo. Más allá de este límite, los errores en el genoma al replicar hacen que el virus sea inútil o no competitivo. Para compensar esto, los virus de ARN a menudo tienen genomas segmentados, el genoma se divide en moléculas más pequeñas, lo que reduce la posibilidad de que un error en un genoma de un solo componente incapacite al genoma completo. Por el contrario, los virus de ADN generalmente tienen genomas más grandes debido a la alta fidelidad de sus enzimas de replicación. Los virus de ADN monocatenario son una excepción a esta regla, ya que las tasas de mutación para estos genomas pueden acercarse al extremo del caso del virus ssRNA.
Mutación genética
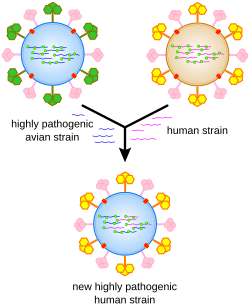
Los virus sufren cambios genéticos por varios mecanismos. Estos incluyen un proceso llamado deriva antigénica donde las bases individuales en el ADN o ARN mutan a otras bases. La mayoría de estas mutaciones puntuales son "silenciosas", no cambian la proteína que codifica el gen, pero otras pueden conferir ventajas evolutivas, como la resistencia a los medicamentos antivirales. El cambio antigénico ocurre cuando hay un cambio importante en el genoma del virus. Esto puede ser el resultado de recombinación o recombinación. Cuando esto sucede con los virus de la influenza, pueden producirse pandemias. Los virus de ARN a menudo existen como cuasiespecies o enjambres de virus de la misma especie pero con secuencias de nucleósidos del genoma ligeramente diferentes. Tales cuasiespecies son un objetivo principal para la selección natural.
Los genomas segmentados confieren ventajas evolutivas; diferentes cepas de un virus con un genoma segmentado pueden mezclar y combinar genes y producir virus de progenie o (descendencia) que tienen características únicas. Esto se llama reasignación o sexo viral .
La recombinación genética es el proceso por el cual una cadena de ADN se rompe y luego se une al extremo de una molécula de ADN diferente. Esto puede ocurrir cuando los virus infectan las células simultáneamente y los estudios de evolución viral han demostrado que la recombinación ha sido desenfrenada en las especies estudiadas. La recombinación es común para los virus de ARN y ADN.
Ciclo de replicación
Las poblaciones virales no crecen a través de la división celular, porque son acelulares. En cambio, usan la maquinaria y el metabolismo de una célula huésped para producir copias múltiples de ellos mismos, y se reúnen en la célula.
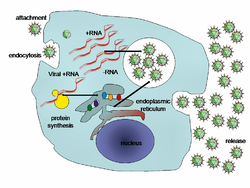
El ciclo de vida de los virus difiere mucho entre las especies, pero hay seis etapas básicas en el ciclo de vida de los virus:
El apego es una unión específica entre las proteínas de la cápside viral y los receptores específicos en la superficie celular del huésped. Esta especificidad determina el rango de host de un virus. Por ejemplo, el VIH infecta un rango limitado de leucocitos humanos. Esto se debe a que su proteína de superficie, gp120, interactúa específicamente con la molécula de CD4, un receptor de quimiocinas, que se encuentra con mayor frecuencia en la superficie de las células T CD4 +. Este mecanismo ha evolucionado para favorecer a aquellos virus que infectan solo las células en las que son capaces de replicarse. La unión al receptor puede inducir a que la proteína de la envoltura viral experimente cambios que produzcan la fusión de membranas virales y celulares, o cambios de las proteínas de la superficie del virus sin envoltura que permiten la entrada del virus.
Penetración sigue a la unión: los viriones ingresan a la célula huésped a través de endocitosis mediada por receptor o fusión de membrana. Esto a menudo se llama entrada viral. La infección de plantas y células fúngicas es diferente de la de las células animales. Las plantas tienen una pared celular rígida hecha de celulosa y hongos de quitina, por lo que la mayoría de los virus pueden ingresar a estas células solo después de un trauma en la pared celular. Casi todos los virus de plantas (como el virus del mosaico del tabaco) también pueden moverse directamente de una célula a otra, en forma de complejos de nucleoproteína monocatenarios, a través de poros llamados plasmodesmata. Las bacterias, como las plantas, tienen paredes celulares fuertes que un virus debe traspasar para infectar la célula. Dado que las paredes de las células bacterianas son mucho más delgadas que las paredes de las células vegetales debido a su tamaño mucho más pequeño,
La desnaturalización es un proceso en el que se elimina la cápside viral: esto puede ser por degradación por enzimas virales o enzimas del huésped o por simple disociación; el resultado final es la liberación del ácido nucleico genómico viral.
La replicación de virus implica principalmente la multiplicación del genoma. La replicación implica la síntesis de ARN mensajero viral (ARNm) de genes "tempranos" (con excepciones para virus de ARN de sentido positivo), síntesis de proteínas virales, posible ensamblaje de proteínas virales, luego replicación del genoma vírico mediado por expresión proteica temprana o reguladora. Esto puede seguirse, para virus complejos con genomas más grandes, por una o más rondas adicionales de síntesis de ARNm: la expresión génica "tardía" es, en general, de proteínas estructurales o de virión.
Ensamblaje : después del autoensamblaje de las partículas de virusmediado por estructuras, amenudo ocurre alguna modificación de las proteínas. En virus como el VIH, esta modificación (a veces llamada maduración) ocurre después de que el virus ha sido liberado de la célula huésped.
Versión - Los virus pueden ser liberados de la célula huésped por lisis, un proceso que mata a la célula al estallar su membrana y la pared celular si está presente: esta es una característica de muchos virus bacterianos y algunos animales. Algunos virus se someten a un ciclo lisogénico en el que el genoma viral se incorpora por recombinación genética a un lugar específico en el cromosoma del huésped. El genoma viral se conoce entonces como un "provirus" o, en el caso de los bacteriófagos, un "profago". Cada vez que el anfitrión se divide, el genoma viral también se replica. El genoma viral es principalmente silencioso dentro del host. En algún momento, el provirus o profago puede dar lugar a un virus activo, que puede lisar las células hospedadoras. Los virus desarrollados (p. Ej., VIH) típicamente se liberan de la célula huésped por gemación. Durante este proceso, el virus adquiere su envoltura, que es una pieza modificada del plasma u otro del huésped,
Replicación del genoma
El material genético dentro de las partículas de virus, y el método por el cual el material se replica, varía considerablemente entre diferentes tipos de virus.
- Virus de ADN
- La replicación del genoma de la mayoría de los virus de ADN tiene lugar en el núcleo de la célula. Si la célula tiene el receptor apropiado en su superficie, estos virus entran en la célula a veces por fusión directa con la membrana celular (p. Ej., Herpesvirus) o, más habitualmente, por endocitosis mediada por receptor. La mayoría de los virus de ADN dependen por completo de la maquinaria de síntesis de ADN y ARN de la célula huésped y de la maquinaria de procesamiento de ARN. Los virus con genomas más grandes pueden codificar mucha de esta maquinaria ellos mismos. En eucariotas, el genoma viral debe cruzar la membrana nuclear de la célula para acceder a esta maquinaria, mientras que en las bacterias solo necesita ingresar a la célula.
- Virus de ARN
- La replicación generalmente tiene lugar en el citoplasma. Los virus de ARN se pueden colocar en cuatro grupos diferentes según sus modos de replicación. La polaridad (si puede o no ser utilizada directamente por los ribosomas para producir proteínas) de los virus de ARN monocatenarios determina en gran medida el mecanismo de replicación; el otro criterio principal es si el material genético es monocatenario o bicatenario. Todos los virus ARN usan sus propias enzimas ARN replicasa para crear copias de sus genomas.
- Invertir la transcripción de virus
- Estos tienen ssRNA ( Retroviridae , Metaviridae , Pseudoviridae ) o dsDNA ( Caulimoviridae y Hepadnaviridae) en sus partículas. Los virus de transcripción reversa con genomas de ARN (retrovirus) usan un intermediario de ADN para replicarse, mientras que aquellos con genomas de ADN (pararetrovirus) usan un ARN intermedio durante la replicación del genoma. Ambos tipos usan una transcriptasa inversa, o la enzima DNA polimerasa dependiente de RNA, para llevar a cabo la conversión de ácido nucleico. Los retrovirus integran el ADN producido por transcripción inversa en el genoma del huésped como un provirus como parte del proceso de replicación; los pararetrovirus no lo hacen, aunque las copias integradas del genoma de pararetrovirus especialmente vegetales pueden dar lugar a virus infecciosos. Son susceptibles a los medicamentos antivirales que inhiben la enzima transcriptasa inversa, por ejemplo, zidovudina y lamivudina. Un ejemplo del primer tipo es el VIH, que es un retrovirus. Ejemplos del segundo tipo son Hepadnaviridae , que incluye el virus de la Hepatitis B
Efectos citopáticos en la célula huésped
El rango de efectos estructurales y bioquímicos que los virus tienen sobre la célula huésped es extenso. Estos se llaman efectos citopáticos . La mayoría de las infecciones de virus eventualmente resultan en la muerte de la célula huésped. Las causas de muerte incluyen lisis celular, alteraciones de la membrana superficial de la célula y apoptosis. A menudo la muerte celular es causada por el cese de sus actividades normales debido a la supresión de proteínas específicas del virus, no todas ellas son componentes de la partícula del virus. La distinción entre citopático e inofensivo es gradual. Algunos virus, como el virus de Epstein-Barr, pueden causar que las células proliferen sin causar malignidad, mientras que otros, como los papilomavirus, son causas establecidas de cáncer.
Infecciones latentes y latentes
Algunos virus no causan cambios aparentes en la célula infectada. Las células en las que el virus está latente e inactivo muestran pocos signos de infección y con frecuencia funcionan normalmente. Esto causa infecciones persistentes y el virus a menudo está inactivo durante muchos meses o años. Este es a menudo el caso con los virus del herpes.
Rango de host
Los virus son, con mucho, las entidades biológicas más abundantes en la Tierra y superan a todos los demás juntos. Infectan todo tipo de vida celular, incluidos animales, plantas, bacterias y hongos. Diferentes tipos de virus pueden infectar solo un rango limitado de hosts y muchos son específicos de cada especie. Algunos, como el virus de la viruela, por ejemplo, pueden infectar solo una especie, en este caso los humanos, y se dice que tienen un rango de hospedadores estrecho. Otros virus, como el virus de la rabia, pueden infectar a diferentes especies de mamíferos y se dice que tienen un amplio rango. Los virus que infectan a las plantas son inofensivos para los animales, y la mayoría de los virus que infectan a otros animales son inofensivos para los humanos. El rango de hospedadores de algunos bacteriófagos se limita a una única cepa de bacterias y se pueden usar para rastrear la fuente de brotes de infecciones mediante un método llamado tipaje de fagos.
Clasificación
La clasificación busca describir la diversidad de virus al nombrarlos y agruparlos sobre la base de similitudes. En 1962, André Lwoff, Robert Horne y Paul Tournier fueron los primeros en desarrollar una clasificación de virus basada en el sistema jerárquico de Linnaean. Esta clasificación basada en el sistema de phylum, clase, orden, familia, género y especie. Los virus se agruparon según sus propiedades compartidas (no las de sus huéspedes) y el tipo de ácido nucleico que forma sus genomas. En 1966, se formó el Comité Internacional de Taxonomía de Virus (ICTV). El sistema propuesto por Lwoff, Horne y Tournier nunca fue totalmente aceptado por el ICTV debido a que los virus de pequeño tamaño del genoma y su alta tasa de mutación dificultan determinar su ascendencia fuera de orden. Como tal,
Clasificación ICTV
El Comité Internacional de Taxonomía de Virus (ICTV) desarrolló el sistema de clasificación actual y escribió pautas que otorgan mayor importancia a ciertas propiedades del virus para mantener la uniformidad familiar. Se ha establecido una taxonomía unificada (un sistema universal para clasificar virus). Solo se ha estudiado una pequeña parte de la diversidad total de virus.
La estructura taxonómica general es la siguiente:
- Orden (- virales )
- Familia (- viridae )
- Subfamilia (- virinae )
- Género ( -virus )
- Especie ( -virus )
- Género ( -virus )
- Subfamilia (- virinae )
- Familia (- viridae )
A partir de 2017, el ICTV ha definido nueve pedidos, 131 familias, 46 subfamilias, 803 géneros y 4,853 especies de virus. Las órdenes son Caudovirales, Herpesvirales, Ligamenvirales, Mononegavirales, Nidovirales, Ortervirales, Picornavirales, Bunyavirales y Tymovirales .
Clasificación de Baltimore

El biólogo ganador del Premio Nobel David Baltimore diseñó el sistema de clasificación de Baltimore. El sistema de clasificación ICTV se usa junto con el sistema de clasificación de Baltimore en la clasificación de virus moderna.
La clasificación de virus de Baltimore se basa en el mecanismo de producción de ARNm. Los virus deben generar ARNm a partir de sus genomas para producir proteínas y replicarse, pero se usan diferentes mecanismos para lograr esto en cada familia de virus. Los genomas virales pueden ser monocatenarios (ss) o bicatenarios (ds), ARN o ADN, y pueden o no usar transcriptasa inversa (RT). Además, los virus ssRNA pueden ser sentido (+) o antisentido (-). Esta clasificación ubica los virus en siete grupos:
- I: virus de dsDNA (por ejemplo, Adenovirus, Herpesvirus, Poxvirus)
- II: ADN de virus de cadena corta (+ cadena o "sentido") de ADN (por ejemplo, parvovirus)
- III: virus dsRNA (p. Ej., Reovirus)
- IV: (+) ARN ssRNA (+ cadena o sentido) ARN (p. Ej. Picornavirus, togavirus)
- V: (-) ARN ssRNA (- cadena o antisentido) RNA (p. Ej. Orthomyxoviruses, Rhabdoviruses)
- VI: ARN ssRNA-RT (+ cadena o sentido) ARN con ADN intermedio en el ciclo de vida (por ejemplo, retrovirus)
- VII: virus dsDNA-RT ADN con ARN intermedio en el ciclo de vida (p. Ej. Hepadnavirus)
Como ejemplo de clasificación viral, el virus varicela zóster (VZV) pertenece al orden Herpesvirales, familia Herpesviridae , subfamilia Alphaherpesvirinae y género Varicellovirus . VZV está en el Grupo I de la Clasificación de Baltimore porque es un virus de dsDNA que no usa transcriptasa inversa.
El conjunto completo de virus en un organismo o hábitat se llama virome; por ejemplo, todos los virus humanos constituyen el viroma humano.
Papel en la enfermedad humana

Los ejemplos de enfermedades humanas comunes causadas por virus incluyen el resfriado común, la influenza, la varicela y el herpes labial. Muchas enfermedades graves como la enfermedad del virus del Ébola, el SIDA, la gripe aviar y el SARS son causadas por virus. La capacidad relativa de los virus para causar enfermedad se describe en términos de virulencia. Se están investigando otras enfermedades para descubrir si tienen un virus como agente causal, como la posible conexión entre el herpesvirus humano 6 (HHV6) y enfermedades neurológicas como la esclerosis múltiple y el síndrome de fatiga crónica. Existe controversia sobre si el bornavirus, que anteriormente se pensaba que causaba enfermedades neurológicas en caballos, podría ser responsable de enfermedades psiquiátricas en humanos.
Los virus tienen diferentes mecanismos por los cuales producen enfermedades en un organismo, que depende en gran medida de la especie viral. Los mecanismos a nivel celular incluyen principalmente la lisis celular, la ruptura abierta y la posterior muerte de la célula. En organismos multicelulares, si mueren suficientes células, todo el organismo comenzará a sufrir los efectos. Aunque los virus causan la interrupción de la homeostasis saludable, lo que resulta en una enfermedad, pueden existir relativamente inofensivos dentro de un organismo. Un ejemplo incluiría la capacidad del virus del herpes simple, que causa el herpes labial, para permanecer en estado latente dentro del cuerpo humano. Esto se llama latencia y es una característica de los virus del herpes, incluido el virus de Epstein-Barr, que causa la fiebre glandular, y el virus de la varicela zoster, que causa la varicela y el herpes zóster. La mayoría de las personas han sido infectadas con al menos uno de estos tipos de virus del herpes. Estos virus latentes a veces pueden ser beneficiosos, ya que la presencia del virus puede aumentar la inmunidad contra patógenos bacterianos, como Yersinia pestis .
Algunos virus pueden causar infecciones de por vida o crónicas, donde los virus continúan reproduciéndose en el cuerpo a pesar de los mecanismos de defensa del huésped. Esto es común en las infecciones por el virus de la hepatitis B y el virus de la hepatitis C. Las personas con infección crónica se conocen como portadores, ya que sirven como reservorios de virus infecciosos. En poblaciones con una alta proporción de portadores, se dice que la enfermedad es endémica.
Epidemiología
La epidemiología viral es la rama de la ciencia médica que se ocupa de la transmisión y el control de las infecciones virales en humanos. La transmisión de virus puede ser vertical, lo que significa de madre a hijo u horizontal, lo que significa de persona a persona. Los ejemplos de transmisión vertical incluyen el virus de la hepatitis B y el VIH, donde el bebé nace ya infectado con el virus. Otro ejemplo, más raro, es el virus de la varicela zóster, que aunque causa infecciones relativamente leves en los seres humanos, puede ser fatal para el feto y el bebé recién nacido.
La transmisión horizontal es el mecanismo más común de propagación de virus en poblaciones. La transmisión puede ocurrir cuando: se intercambian fluidos corporales durante la actividad sexual, por ejemplo, VIH; la sangre se intercambia por transfusión contaminada o por intercambio de agujas, por ejemplo, hepatitis C; intercambio de saliva por vía oral, p. ej., virus de Epstein-Barr; se ingiere alimento o agua contaminados, por ejemplo, norovirus; aerosoles que contienen viriones son inhalados, por ejemplo, virus de la gripe; y vectores de insectos tales como mosquitos que penetran en la piel de un huésped, por ejemplo, el dengue. La tasa o velocidad de transmisión de las infecciones virales depende de factores que incluyen la densidad de población, el número de individuos susceptibles (es decir, aquellos que no son inmunes), la calidad de la atención médica y el clima.
La epidemiología se usa para romper la cadena de infección en las poblaciones durante los brotes de enfermedades virales. Se utilizan medidas de control que se basan en el conocimiento de cómo se transmite el virus. Es importante encontrar la fuente o fuentes del brote e identificar el virus. Una vez que el virus ha sido identificado, la cadena de transmisión a veces puede romperse con vacunas. Cuando las vacunas no están disponibles, el saneamiento y la desinfección pueden ser efectivos. A menudo, las personas infectadas están aisladas del resto de la comunidad, y las que han estado expuestas al virus se ponen en cuarentena. Para controlar el brote de fiebre aftosa en el ganado en Gran Bretaña en 2001, se sacrificaron miles de cabezas de ganado. La mayoría de las infecciones virales de humanos y otros animales tienen períodos de incubación durante los cuales la infección no causa signos o síntomas. Los períodos de incubación de las enfermedades virales oscilan entre algunos días y semanas, pero son conocidos por la mayoría de las infecciones. Algo superpuesto, pero principalmente después del período de incubación, hay un período de comunicabilidad: un momento en que una persona o animal infectado es contagioso y puede infectar a otra persona o animal. Esto también es conocido por muchas infecciones virales, y el conocimiento de la duración de ambos períodos es importante para controlar los brotes. Cuando los brotes causan una proporción inusualmente alta de casos en una población, comunidad o región, se llaman epidemias. Si los brotes se propagan por todo el mundo, se llaman pandemias. hay un período de comunicabilidad: un momento en que una persona o animal infectado es contagioso y puede infectar a otra persona o animal. Esto también es conocido por muchas infecciones virales, y el conocimiento de la duración de ambos períodos es importante para controlar los brotes. Cuando los brotes causan una proporción inusualmente alta de casos en una población, comunidad o región, se llaman epidemias. Si los brotes se propagan por todo el mundo, se llaman pandemias. hay un período de comunicabilidad: un momento en que una persona o animal infectado es contagioso y puede infectar a otra persona o animal. Esto también es conocido por muchas infecciones virales, y el conocimiento de la duración de ambos períodos es importante para controlar los brotes. Cuando los brotes causan una proporción inusualmente alta de casos en una población, comunidad o región, se llaman epidemias. Si los brotes se propagan por todo el mundo, se llaman pandemias.
Epidemias y pandemias
Las poblaciones de nativos americanos fueron devastadas por enfermedades contagiosas, en particular, la viruela, traídas a las Américas por colonos europeos. No está claro cuántos nativos americanos murieron a causa de enfermedades extranjeras después de la llegada de Colón a las Américas, pero se estima que el número se acerca al 70% de la población indígena. El daño causado por esta enfermedad ayudó significativamente a los intentos europeos de desplazar y conquistar a la población nativa.
Una pandemia es una epidemia mundial. La pandemia de gripe de 1918, que duró hasta 1919, fue una pandemia de influenza de categoría 5 causada por un virus de influenza A inusualmente grave y mortal. Las víctimas a menudo eran adultos jóvenes sanos, a diferencia de la mayoría de los brotes de influenza, que afectan predominantemente a pacientes menores, ancianos o debilitados. Las estimaciones anteriores dicen que mató a 40-50 millones de personas, mientras que investigaciones más recientes sugieren que pudo haber matado a hasta 100 millones de personas, o el 5% de la población mundial en 1918.
La mayoría de los investigadores creen que el VIH se originó en el África subsahariana durante el siglo XX; ahora es una pandemia, con un estimado de 38.6 millones de personas que ahora viven con la enfermedad en todo el mundo. El Programa Conjunto de las Naciones Unidas sobre el VIH / SIDA (ONUSIDA) y la Organización Mundial de la Salud (OMS) estiman que el SIDA ha matado a más de 25 millones de personas desde que se reconoció por primera vez el 5 de junio de 1981, convirtiéndola en una de las epidemias más destructivas historia. En 2007 hubo 2,7 millones de nuevas infecciones por VIH y 2 millones de muertes relacionadas con el VIH.


Varios agentes patógenos virales altamente letales son miembros de los Filoviridae . Los filovirus son virus filamentosos que causan fiebre hemorrágica viral e incluyen ebolavirus y marburgvirus. El virus de Marburg, descubierto por primera vez en 1967, atrajo la atención de la prensa en abril de 2005 por un brote en Angola. La enfermedad del virus del Ébola también ha causado brotes intermitentes con altas tasas de mortalidad desde 1976 cuando se identificó por primera vez. La peor y la más reciente es la epidemia de África occidental.
Cáncer
Los virus son una causa establecida de cáncer en humanos y otras especies. Los cánceres virales ocurren solo en una minoría de personas infectadas (o animales). Los virus del cáncer provienen de una variedad de familias de virus, incluidos los virus de ARN y ADN, por lo que no existe un solo tipo de "oncovirus" (un término obsoleto usado originalmente para la transformación aguda de retrovirus). El desarrollo del cáncer está determinado por diversos factores, como la inmunidad del huésped y las mutaciones en el huésped. Los virus aceptados como causantes del cáncer humano incluyen algunos genotipos del virus del papiloma humano, virus de la hepatitis B, virus de la hepatitis C, virus de Epstein-Barr y sarcoma de Kaposi. herpesvirus asociado y virus T-linfotrópico humano. El virus del cáncer humano descubierto más recientemente es un poliomavirus (poliomavirus de células de Merkel) que causa la mayoría de los casos de una forma rara de cáncer de piel llamado carcinoma de células de Merkel. Los virus de hepatitis pueden convertirse en una infección viral crónica que conduce al cáncer de hígado. La infección por el virus linfotrópico T humano puede conducir a la paraparesia espástica tropical y a la leucemia de células T adultas. Los virus del papiloma humano son una causa establecida de cánceres de cuello uterino, piel, ano y pene. Dentro de Herpesviridae , el virus del herpes asociado al sarcoma de Kaposi causa sarcoma de Kaposi y linfoma de la cavidad corporal, y el virus de Epstein-Barr causa el linfoma de Burkitt, el linfoma de Hodgkin, el trastorno linfoproliferativo B y el carcinoma nasofaríngeo. El poliomavirus de células de Merkel está estrechamente relacionado con el SV40 y los poliomavirus de ratón que se han utilizado como modelos animales para los virus del cáncer durante más de 50 años.
Mecanismos de defensa del huésped
La primera línea de defensa del cuerpo contra los virus es el sistema inmune innato. Esto comprende células y otros mecanismos que defienden al huésped de la infección de una manera no específica. Esto significa que las células del sistema innato reconocen y responden a los patógenos de una manera genérica, pero, a diferencia del sistema inmune adaptativo, no confiere inmunidad duradera o protectora al huésped.
La interferencia de ARN es una defensa innata importante contra los virus. Muchos virus tienen una estrategia de replicación que implica ARN bicatenario (dsRNA). Cuando un virus de este tipo infecta a una célula, libera su molécula o moléculas de ARN, que se unen inmediatamente a un complejo de proteína llamado dicer que corta el ARN en pedazos más pequeños. Se activa una ruta bioquímica, el complejo RISC, que garantiza la supervivencia celular al degradar el ARNm viral. Los rotavirus han evolucionado para evitar este mecanismo de defensa al no desenredarse por completo dentro de la célula y liberar ARNm recién producido a través de los poros en la cápside interna de la partícula. Su dsRNA genómico permanece protegido dentro del núcleo del virión.
Cuando el sistema inmune adaptativo de un vertebrado se encuentra con un virus, produce anticuerpos específicos que se unen al virus y, a menudo, lo vuelven no infeccioso. Esto se llama inmunidad humoral. Dos tipos de anticuerpos son importantes. La primera, llamada IgM, es altamente efectiva para neutralizar virus pero es producida por las células del sistema inmune solo por unas pocas semanas. El segundo, llamado IgG, se produce indefinidamente. La presencia de IgM en la sangre del huésped se usa para detectar infección aguda, mientras que IgG indica una infección en el pasado. El anticuerpo IgG se mide cuando se realizan pruebas de inmunidad.
Los anticuerpos pueden seguir siendo un mecanismo de defensa eficaz incluso después de que los virus hayan logrado ingresar a la célula huésped. Una proteína que está en las células, llamada TRIM21, puede unirse a los anticuerpos en la superficie de la partícula del virus. Esto prepara la posterior destrucción del virus por las enzimas del sistema proteosómico de la célula.
Una segunda defensa de los vertebrados contra los virus se llama inmunidad mediada por células e involucra a las células inmunes conocidas como células T. Las células del cuerpo muestran constantemente fragmentos cortos de sus proteínas en la superficie de la célula y, si una célula T reconoce allí un fragmento viral sospechoso, la célula huésped es destruida por las células T asesinas y las células T específicas del virus proliferan. Las células como el macrófago son especialistas en esta presentación de antígeno. La producción de interferón es un importante mecanismo de defensa del huésped. Esta es una hormona producida por el cuerpo cuando hay virus. Su papel en la inmunidad es complejo; finalmente impide que los virus se reproduzcan al matar a la célula infectada y sus vecinos cercanos.
No todas las infecciones de virus producen una respuesta inmune protectora de esta manera. El VIH evade el sistema inmune cambiando constantemente la secuencia de aminoácidos de las proteínas en la superficie del virión. Esto se conoce como "mutación de escape" ya que los epítopos virales escapan al reconocimiento por la respuesta inmune del huésped. Estos virus persistentes evaden el control inmune por secuestro, bloqueo de la presentación del antígeno, resistencia a las citoquinas, evasión de las actividades de las células asesinas naturales, escape de la apoptosis y cambio antigénico. Otros virus, llamados virus neurotrópicos , se diseminan por la diseminación de los nervios donde el sistema inmunitario puede ser incapaz de alcanzarlos.
Prevención y tratamiento
Debido a que los virus utilizan vías metabólicas vitales dentro de las células del huésped para replicarse, son difíciles de eliminar sin usar drogas que causan efectos tóxicos a las células hospedadoras en general. Los enfoques médicos más efectivos para las enfermedades virales son las vacunas para proporcionar inmunidad a la infección y los medicamentos antivirales que interfieren selectivamente con la replicación viral.
Vacunas
La vacunación es una forma barata y efectiva de prevenir infecciones por virus. Las vacunas se usaron para prevenir infecciones virales mucho antes del descubrimiento de los virus reales. Su uso ha resultado en una disminución dramática en la morbilidad (enfermedad) y la mortalidad (muerte) asociada con infecciones virales como la polio, el sarampión, las paperas y la rubéola. Las infecciones de viruela han sido erradicadas. Las vacunas están disponibles para prevenir más de trece infecciones virales de los seres humanos, y más se utilizan para prevenir las infecciones virales de los animales. Las vacunas pueden consistir en virus vivos atenuados o muertos, o proteínas virales (antígenos). Las vacunas vivas contienen formas debilitadas del virus, que no causan la enfermedad pero, sin embargo, confieren inmunidad. Dichos virus se llaman atenuados. Las vacunas vivas pueden ser peligrosas cuando se administran a personas con una inmunidad débil (que se describen como inmunocomprometidas), porque en estas personas, el virus debilitado puede causar la enfermedad original. Las técnicas de biotecnología e ingeniería genética se utilizan para producir vacunas de subunidades. Estas vacunas usan solo las proteínas de la cápside del virus. La vacuna contra la hepatitis B es un ejemplo de este tipo de vacuna. Las vacunas de subunidades son seguras para pacientes inmunocomprometidos porque no pueden causar la enfermedad. La vacuna contra el virus de la fiebre amarilla, una cepa atenuada en vivo llamada 17D, es probablemente la vacuna más segura y efectiva jamás generada. La vacuna contra la hepatitis B es un ejemplo de este tipo de vacuna. Las vacunas de subunidades son seguras para pacientes inmunocomprometidos porque no pueden causar la enfermedad. La vacuna contra el virus de la fiebre amarilla, una cepa atenuada en vivo llamada 17D, es probablemente la vacuna más segura y efectiva jamás generada. La vacuna contra la hepatitis B es un ejemplo de este tipo de vacuna. Las vacunas de subunidades son seguras para pacientes inmunocomprometidos porque no pueden causar la enfermedad. La vacuna contra el virus de la fiebre amarilla, una cepa atenuada en vivo llamada 17D, es probablemente la vacuna más segura y efectiva jamás generada.
Drogas antivirales

Los medicamentos antivirales a menudo son análogos de nucleósidos (bloques de construcción de ADN falsos), que los virus incorporan por error en sus genomas durante la replicación. El ciclo de vida del virus se detiene porque el ADN recién sintetizado está inactivo. Esto se debe a que estos análogos carecen de los grupos hidroxilo, que, junto con los átomos de fósforo, se unen para formar la "columna vertebral" fuerte de la molécula de ADN. Esto se llama terminación de la cadena de ADN. Ejemplos de análogos de nucleósidos son aciclovir para infecciones por virus Herpes simplex y lamivudina para infecciones por VIH y virus de hepatitis B. Aciclovir es uno de los medicamentos antivirales más antiguos y recetados con mayor frecuencia. Otros medicamentos antivirales en uso se dirigen a diferentes etapas del ciclo de vida viral. El VIH depende de una enzima proteolítica llamada proteasa del VIH-1 para que se vuelva completamente infecciosa.
La hepatitis C es causada por un virus de ARN. En el 80% de las personas infectadas, la enfermedad es crónica, y sin tratamiento, están infectadas por el resto de sus vidas. Ahora hay un tratamiento efectivo que usa el fármaco análogo de nucleósido ribavirina combinado con interferón. Se ha desarrollado el tratamiento de los portadores crónicos del virus de la hepatitis B utilizando una estrategia similar usando lamivudina.
Infección en otras especies
Los virus infectan toda la vida celular y, aunque los virus se producen universalmente, cada especie celular tiene su propio rango específico que a menudo infecta solo a esa especie. Algunos virus, llamados satélites, pueden replicarse solo dentro de las células que ya han sido infectadas por otro virus.
Virus de animales
Los virus son importantes patógenos del ganado. Las enfermedades como la fiebre aftosa y la lengua azul son causadas por virus. Los animales de compañía, como gatos, perros y caballos, si no están vacunados, son susceptibles a infecciones virales graves. El parvovirus canino es causado por un pequeño virus de ADN y las infecciones a menudo son fatales en los cachorros. Como todos los invertebrados, la abeja melífera es susceptible a muchas infecciones virales. La mayoría de los virus coexisten inofensivamente en su huésped y no causan signos o síntomas de enfermedad.
Virus de plantas

Hay muchos tipos de virus de plantas, pero a menudo solo causan una pérdida de rendimiento, y no es económicamente viable tratar de controlarlos. Los virus de las plantas a menudo se propagan de una planta a otra por organismos, conocidos como vectores . Estos son normalmente insectos, pero algunos hongos, gusanos nematodos y organismos unicelulares han demostrado ser vectores. Cuando el control de las infecciones por virus de plantas se considera económico, para las frutas perennes, por ejemplo, los esfuerzos se concentran en matar los vectores y eliminar los huéspedes alternativos, como las malas hierbas. Los virus de plantas no pueden infectar a humanos y otros animales porque pueden reproducirse solo en las células vegetales vivas.
Las plantas tienen mecanismos de defensa elaborados y efectivos contra los virus. Una de las más efectivas es la presencia de los llamados genes de resistencia (R). Cada gen R confiere resistencia a un virus particular activando áreas localizadas de muerte celular alrededor de la célula infectada, que a menudo se pueden ver a simple vista como puntos grandes. Esto evita que la infección se propague. La interferencia de ARN también es una defensa efectiva en las plantas. Cuando se infectan, las plantas a menudo producen desinfectantes naturales que matan a los virus, como el ácido salicílico, el óxido nítrico y las moléculas reactivas de oxígeno.
Las partículas de virus de plantas o partículas similares a virus (VLP) tienen aplicaciones tanto en biotecnología como en nanotecnología. Las cápsides de la mayoría de los virus de plantas son estructuras simples y robustas y pueden producirse en grandes cantidades ya sea por la infección de las plantas o por la expresión en una variedad de sistemas heterólogos. Las partículas de virus de plantas pueden modificarse genéticamente y químicamente para encapsular material extraño y pueden incorporarse en estructuras supramoleculares para su uso en biotecnología.
Virus bacterianos

Los bacteriófagos son un grupo común y diverso de virus y son la entidad biológica más abundante en ambientes acuáticos: hay hasta 10 veces más de estos virus en los océanos que bacterias, alcanzando niveles de 250,000,000 de bacteriófagos por mililitro de agua de mar. Estos virus infectan bacterias específicas uniéndose a las moléculas receptoras de superficie y luego ingresando a la célula. En un corto período de tiempo, en algunos casos solo minutos, la polimerasa bacteriana comienza a traducir el ARNm viral en proteína. Estas proteínas se convierten en viriones nuevos dentro de la célula, proteínas auxiliares, que ayudan al ensamblaje de nuevos viriones o proteínas involucradas en la lisis celular. Las enzimas virales ayudan a la descomposición de la membrana celular y, en el caso del fago T4, en poco más de veinte minutos después de la inyección podrían liberarse más de trescientos fagos.
La principal forma en que las bacterias se defienden de los bacteriófagos es produciendo enzimas que destruyen el ADN extraño. Estas enzimas, llamadas endonucleasas de restricción, cortan el ADN viral que los bacteriófagos inyectan en las células bacterianas. Las bacterias también contienen un sistema que utiliza secuencias CRISPR para retener fragmentos de los genomas de los virus con los que las bacterias han entrado en contacto en el pasado, lo que les permite bloquear la replicación del virus a través de una forma de interferencia de ARN. Este sistema genético proporciona a las bacterias inmunidad adquirida a la infección.
Virus de Archaeal
Algunos virus se replican dentro de las arqueas: estos son virus de ADN bicatenario con formas inusuales ya veces únicas. Estos virus han sido estudiados con mayor detalle en las arqueas termófilas, particularmente en los órdenes Sulfolobales y Thermoproteales. Las defensas contra estos virus implican interferencia de ARN a partir de secuencias de ADN repetitivas dentro de genomas de arqueas que están relacionadas con los genes de los virus. La mayoría de las arqueas tienen sistemas CRISPR-Cas como una defensa adaptativa contra los virus. Estos permiten a las arqueas retener secciones de ADN viral, que luego se usan para atacar y eliminar las infecciones subsiguientes del virus usando un proceso similar a la interferencia del ARN.
Papel en los ecosistemas acuáticos
Una cucharadita de agua de mar (~ 5 ml) contiene alrededor de 50 millones de virus, que contienen una enorme diversidad genética. La mayoría de estos son virus que infectan bacterias heterotróficas "bacteriófagos" y cianobacterias "cianófagos", que son inofensivos para las plantas y los animales, y son esenciales para la regulación de los ecosistemas marinos y de agua dulce; también son importantes agentes de mortalidad del fitoplancton, la base de la cadena alimenticia en ambientes acuáticos. Infectan y destruyen las bacterias en las comunidades microbianas acuáticas, y son uno de los mecanismos más importantes para reciclar el carbono y el ciclo de nutrientes en los ambientes marinos. Las moléculas orgánicas liberadas de las células bacterianas muertas estimulan el crecimiento fresco de bacterias y algas, en un proceso conocido como derivación viral. En particular, Se ha demostrado que la lisis de las bacterias mediante virus mejora el ciclo del nitrógeno y estimula el crecimiento del fitoplancton. La actividad viral también puede afectar a la bomba biológica, el proceso mediante el cual el carbono se secuestra en las profundidades del océano.
Los microorganismos constituyen más del 90% de la biomasa en el mar. Se estima que los virus matan aproximadamente el 20% de esta biomasa cada día y que hay de 10 a 15 veces más virus en los océanos que bacterias y arqueas. Los virus también son los principales agentes responsables de la destrucción del fitoplancton, incluidas las floraciones de algas nocivas. La cantidad de virus en los océanos disminuye aún más lejos de la costa y más profundamente en el agua, donde hay menos organismos hospedadores.
En enero de 2018, los científicos informaron, por primera vez, que los 800 millones de virus, principalmente de origen marino, se depositan todos los días de la Tierra 's atmósfera en cada metro cuadrado de la superficie del planeta, como el resultado de una corriente atmosférica global de los virus , circulando por encima del sistema meteorológico, pero por debajo de la altura de los viajes habituales en avión, distribuyendo virus por todo el planeta.
Al igual que cualquier organismo, los mamíferos marinos son susceptibles a las infecciones virales. En 1988 y 2002, miles de focas portuarias fueron asesinadas en Europa por el virus del moquillo fúngico. Muchos otros virus, incluidos calicivirus, herpesvirus, adenovirus y parvovirus, circulan en poblaciones de mamíferos marinos.
Papel en la evolución
Los virus son un medio natural importante para transferir genes entre diferentes especies, lo que aumenta la diversidad genética e impulsa la evolución. Se cree que los virus jugaron un papel central en la evolución temprana, antes de la diversificación de bacterias, arqueas y eucariotas, en el momento del último ancestro común universal de la vida en la Tierra. Los virus siguen siendo uno de los mayores reservorios de diversidad genética inexplorada en la Tierra.
Aplicaciones
Ciencias de la vida y medicina

Los virus son importantes para el estudio de la biología molecular y celular ya que proporcionan sistemas simples que pueden usarse para manipular e investigar las funciones de las células. El estudio y el uso de virus han proporcionado información valiosa sobre aspectos de la biología celular. Por ejemplo, los virus han sido útiles en el estudio de la genética y han ayudado a nuestra comprensión de los mecanismos básicos de la genética molecular, como la replicación del ADN, la transcripción, el procesamiento del ARN, la traducción, el transporte de proteínas y la inmunología.
Los genetistas a menudo usan virus como vectores para introducir genes en las células que están estudiando. Esto es útil para hacer que la célula produzca una sustancia extraña, o para estudiar el efecto de la introducción de un nuevo gen en el genoma. De manera similar, la viroterapia utiliza virus como vectores para tratar diversas enfermedades, ya que pueden dirigirse específicamente a las células y al ADN. Muestra un uso prometedor en el tratamiento del cáncer y en la terapia génica. Los científicos de Europa del Este han utilizado la terapia con fagos como una alternativa a los antibióticos durante algún tiempo, y el interés en este enfoque está aumentando, debido al alto nivel de resistencia a los antibióticos que ahora se encuentra en algunas bacterias patógenas. La expresión de proteínas heterólogas por virus es la base de varios procesos de fabricación que actualmente se utilizan para la producción de diversas proteínas, como antígenos de vacunas y anticuerpos. Los procesos industriales se han desarrollado recientemente utilizando vectores virales y varias proteínas farmacéuticas se encuentran actualmente en ensayos preclínicos y clínicos.
Viroterapia
La viroterapia implica el uso de virus genéticamente modificados para tratar enfermedades. Los científicos han modificado los virus para que se reproduzcan en las células cancerígenas y las destruyan, pero no infecten las células sanas. Talimogene laherparepvec (T-VEC), por ejemplo, es un virus herpes simplex modificado que ha tenido un gen, que es necesario para que los virus se repliquen en células sanas, eliminado y reemplazado por un gen humano (GM-CSF) que estimula la inmunidad. Cuando este virus infecta las células cancerígenas, las destruye y, al hacerlo, la presencia del gen GM-CSF atrae a las células dendríticas de los tejidos circundantes del cuerpo. Las células dendríticas procesan las células cancerosas muertas y presentan componentes de ellas en otras células del sistema inmune. Habiendo completado exitosos ensayos clínicos,
Ciencia de materiales y nanotecnología
Las tendencias actuales en nanotecnología prometen hacer un uso mucho más versátil de los virus. Desde el punto de vista de un científico de materiales, los virus pueden considerarse nanopartículas orgánicas. Su superficie lleva herramientas específicas diseñadas para cruzar las barreras de sus células anfitrionas. El tamaño y la forma de los virus, y el número y la naturaleza de los grupos funcionales en su superficie, están definidos con precisión. Como tal, los virus se usan comúnmente en la ciencia de materiales como andamios para modificaciones de superficie unidas covalentemente. Una cualidad particular de los virus es que pueden ser adaptados por la evolución dirigida. Las poderosas técnicas desarrolladas por las ciencias de la vida se están convirtiendo en la base de los enfoques de ingeniería hacia los nanomateriales, abriendo una amplia gama de aplicaciones que van más allá de la biología y la medicina.
Debido a su tamaño, forma y estructuras químicas bien definidas, los virus se han utilizado como plantillas para organizar los materiales a nanoescala. Ejemplos recientes incluyen trabajos en el Naval Research Laboratory en Washington, DC, usando partículas del virus del mosaico de Cowpea (CPMV) para amplificar señales en sensores basados en microarrays de ADN. En esta aplicación, las partículas de virus separan los colorantes fluorescentes utilizados para la señalización para evitar la formación de dímeros no fluorescentes que actúan como atenuadores. Otro ejemplo es el uso de CPMV como una placa de prueba a nanoescala para electrónica molecular.
Virus sintéticos
Muchos virus se pueden sintetizar de novo ("desde cero") y el primer virus sintético se creó en 2002. Aunque es un concepto erróneo, no se sintetiza el virus real, sino su genoma de ADN (en el caso de un ADN) virus), o una copia de cDNA de su genoma (en el caso de virus de ARN). Para muchas familias de virus, el ADN o ARN sintético desnudo (una vez reconvertido enzimáticamente desde el ADNc sintético) es infeccioso cuando se introduce en una célula. Es decir, contienen toda la información necesaria para producir nuevos virus. Esta tecnología ahora se usa para investigar nuevas estrategias de vacunas. La capacidad de sintetizar virus tiene consecuencias de gran alcance, ya que los virus ya no se pueden considerar extintos, siempre que se conozca la información de su secuencia de genoma y se disponga de células permisivas. A partir de noviembre de 2017.
Armas
La capacidad de los virus para causar epidemias devastadoras en las sociedades humanas ha llevado a la preocupación de que los virus puedan ser armados para la guerra biológica. Surgió una mayor preocupación por la exitosa recreación del infame virus de la influenza de 1918 en un laboratorio. El virus de la viruela devastó numerosas sociedades a lo largo de la historia antes de su erradicación. Solo hay dos centros en el mundo que están autorizados por la OMS para mantener existencias de virus de la viruela: el Centro de Investigación Estatal de Virología y Biotecnología VECTOR en Rusia y los Centros para el Control y la Prevención de Enfermedades en los Estados Unidos. El temor de que pueda ser usado como arma puede no ser totalmente infundado. Como la vacuna para la viruela a veces tiene efectos secundarios graves, ya no se usa de manera rutinaria en ningún país.
Obtenido de: https://en.wikipedia.org/wiki/Virus