Abiogénesis
Definición

La abiogénesis , o informalmente el origen de la vida , es el proceso natural por el cual la vida surge de la materia no viva, como los compuestos orgánicos simples. La transición de entidades no vivientes a entidades vivientes no fue un evento único, sino un proceso gradual de complejidad creciente.
Los investigadores estudian la abiogénesis a través de una combinación de biología molecular, paleontología, astrobiología y bioquímica, y su objetivo es determinar cómo las reacciones químicas previas a la vida dieron lugar a la vida. El estudio de la abiogénesis puede ser geofísico, químico o biológico, con enfoques más recientes que intentan una síntesis de los tres, ya que la vida surgió en condiciones que son sorprendentemente diferentes de las de la Tierra en la actualidad. La vida funciona a través de la química especializada del carbono y el agua y se basa principalmente en cuatro familias clave de productos químicos: lípidos (paredes de células grasas), carbohidratos (azúcares, celulosa), aminoácidos (metabolismo de proteínas) y ácidos nucleicos (ADN autoreplicante y ARN). Cualquier teoría exitosa de la abiogénesis debe explicar los orígenes y las interacciones de estas clases de moléculas. Muchos enfoques de la abiogénesis investigan cómo las moléculas autorreplicantes, o sus componentes, llegaron a existir. Los investigadores generalmente piensan que la vida actual en la Tierra desciende de un mundo de ARN, aunque la vida basada en ARN puede no haber sido la primera vida que existió.
El clásico experimento de Miller-Urey de 1952 y una investigación similar demostraron que la mayoría de los aminoácidos, los constituyentes químicos de las proteínas utilizadas en todos los organismos vivos, pueden sintetizarse a partir de compuestos inorgánicos en condiciones destinadas a reproducir los de la Tierra primitiva. Los científicos han propuesto varias fuentes externas de energía que pueden haber desencadenado estas reacciones, incluidos los rayos y la radiación. Otros enfoques (hipótesis del "metabolismo primero") se centran en la comprensión de cómo la catálisis en los sistemas químicos en la Tierra primitiva podría haber proporcionado las moléculas precursoras necesarias para la autorreplicación. Las moléculas orgánicas complejas se producen en el Sistema Solar y en el espacio interestelar, y estas moléculas pueden haber proporcionado material de partida para el desarrollo de la vida en la Tierra.
La bioquímica de la vida puede haber comenzado poco después del Big Bang, hace 13.800 millones de años, durante una época habitable cuando la edad del universo era de solo 10 a 17 millones de años. La hipótesis de la panspermia sugiere que la vida microscópica se distribuyó a la Tierra primitiva por el polvo espacial, los meteoritos, los asteroides y otros cuerpos pequeños del Sistema Solar y que la vida puede existir en todo el universo. La hipótesis de la panspermia propone que la vida se originó fuera de la Tierra, pero no explica definitivamente su origen.
No obstante, la Tierra sigue siendo el único lugar en el universo que se sabe que alberga vida, y la evidencia fósil de la Tierra informa la mayoría de los estudios de la abiogénesis. La edad de la Tierra es de unos 4.540 millones de años; la primera evidencia indiscutible de vida en la Tierra data de hace al menos 3.500 millones de años, y posiblemente tan temprano como la Era Eoarcheana (hace entre 3.600 y 4.000 millones de años), luego de que la corteza geológica comenzara a solidificarse siguiendo el Haón Eón fundido. En mayo de 2017, los científicos encontraron evidencias de vida temprana en tierra en geiseritas de 3.48 billones de años y otros depósitos minerales relacionados (a menudo encontrados alrededor de fuentes termales y géiseres) descubiertos en el Craton Pilbara de Australia Occidental. Sin embargo, varios descubrimientos sugieren que la vida puede haber aparecido en la Tierra incluso antes. A partir de 2017, Los microfósiles dentro de precipitados de ventilación hidrotermal de 3.77 a 4.28 mil millones de años hallados en Quebec, Canadá, pueden ser el registro más antiguo de la vida en la Tierra, sugiriendo que la vida comenzó poco después de la formación oceánica hace 4.400 millones de años. Según el biólogo Stephen Blair Hedges, "si la vida surgiera relativamente rápido en la Tierra ... entonces podría ser común en el universo".
Las primeras condiciones geofísicas en la Tierra
Cronología de vida
-4500 -
-
-4000 -
-
-3500 -
-
-3000 -
-
-2500 -
-
-2000 -
-
-1500 -
-
-1000 -
-
-500 -
-
0 -
Escala del eje: millones de años
Los océanos pueden haber aparecido primero en Hadean Eon, tan pronto como doscientos millones de años (200 Ma) después de que se formó la Tierra, en un ambiente cálido de 100 ° C (212 ° F) y el pH de aproximadamente 5.8 se elevó rápidamente hacia neutral. Esto se ha visto respaldado por la datación de 4.404 cristales de zircón Ga-old de cuarcita metamorfoseada del Monte Narryer en el oeste de Australia Jack Hills de Pilbara, que son evidencia de que los océanos y la corteza continental existían dentro de los 150 Ma de la formación de la Tierra. A pesar del probable volcanismo y la existencia de muchas "plaquetas" tectónicas más pequeñas, se ha sugerido que entre 4.4 y 4.3 Ga (mil millones de años), la Tierra era un mundo acuático, con poca o ninguna corteza continental, una atmósfera extremadamente turbulenta y un hidrosfera sujeta a intensa luz ultravioleta (UV), desde una etapa T Tauri Sol,
El entorno hadeano habría sido muy peligroso para la vida moderna. Las colisiones frecuentes con objetos grandes, de hasta 500 kilómetros de diámetro, habrían sido suficientes para esterilizar el planeta y vaporizar el océano a los pocos meses de impacto, con el vapor caliente mezclado con vapor de roca convirtiéndose en nubes de gran altitud que cubre el planeta Después de unos meses, la altura de estas nubes habría comenzado a disminuir, pero la base de la nube aún se habría elevado durante los próximos mil años. Después de eso, habría comenzado a llover a baja altura. Durante otros dos mil años, las lluvias habrían reducido lentamente la altura de las nubes, devolviendo los océanos a su profundidad original solo 3,000 años después del evento de impacto.
La primera evidencia biológica de la vida
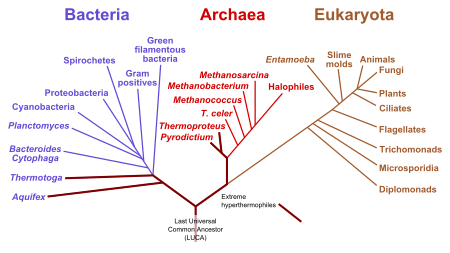
La ubicación más comúnmente aceptada de la raíz del árbol de la vida es entre un dominio monofilético Bacteria y un clado formado por Archaea y Eukaryota de lo que se conoce como el "árbol de la vida tradicional" basado en varios estudios moleculares que comienzan con C. Woese . Una minoría muy pequeña de estudios ha concluido de manera diferente, a saber, que la raíz está en el Dominio de Bacterias, ya sea en el filo Firmicutes o que el phylum Chloroflexi es basal a un clado con Archaea + Eukaryotes y el resto de Bacterias según lo propuesto por Thomas Cavalier- Herrero. Más recientemente, Peter Ward ha establecido una visión alternativa que está arraigada en la síntesis abiótica de ARN que se encerra dentro de una cápsula y luego crea réplicas de ARN ribozima. Se propone que esto se bifurca entre Dominion Ribosa (vida del ARN),

La vida más antigua en la Tierra existió hace más de 3.5 billones de años, durante la Era Eoarcheana cuando la corteza suficiente se había solidificado después del Haón Eón fundido. La evidencia física más temprana hasta ahora encontrada consiste en microfósiles en el Nuvvuagittuq Greenstone Belt del norte de Quebec, en rocas "formadas con bandas de hierro" de al menos 3,77 billones y posiblemente 4,28 billones de años. Este hallazgo sugiere que hubo un desarrollo casi instantáneo de la vida después de la formación de los océanos. Se observó que la estructura de los microbios era similar a la que se encuentra cerca de las fuentes hidrotermales en la era moderna, y apoyó la hipótesis de que la abiogénesis comenzó cerca de los respiraderos hidrotermales.
También es digno de mención el grafito biogénico en rocas metasedimentarias de 3.700 millones de años del sudoeste de Groenlandia y los fósiles de esteras microbianas hallados en areniscas de 3.48 billones de años del oeste de Australia. La evidencia de vida temprana en las rocas de la isla Akilia, cerca del cinturón supracrustal de Isua, en el suroeste de Groenlandia, data de hace 3.700 millones de años y ha mostrado isótopos de carbono biogénicos. En otras partes del cinturón supracrustal de Isua, las inclusiones de grafito atrapadas en los cristales de granate están conectadas a los otros elementos de la vida: oxígeno, nitrógeno y posiblemente fósforo en forma de fosfato, lo que proporciona más evidencia de la vida hace 3.700 millones de años. En Strelley Pool, en la región de Pilbara en el oeste de Australia, se encontraron evidencias convincentes de vida temprana en arenisca con pirita en una playa fosilizada, que mostraba células tubulares redondeadas que oxidaban el azufre por fotosíntesis en ausencia de oxígeno. La investigación adicional sobre zircones de Australia Occidental en 2015 sugirió que la vida probablemente existió en la Tierra hace al menos 4,1 billones de años.
Tradicionalmente se pensaba que durante el período comprendido entre 4,28 y 3,8 Ga, los cambios en las órbitas de los planetas gigantes pudieron haber causado un intenso bombardeo de asteroides y cometas que marcaron la Luna y otros planetas interiores (Mercurio, Marte y, presumiblemente, la Tierra y Venus). Esto probablemente habría esterilizado repetidamente el planeta, la vida apareció antes de ese momento. Geológicamente, la Tierra hadeana habría sido mucho más activa que en cualquier otro momento de su historia. Los estudios de meteoritos sugieren que los isótopos radiactivos como el aluminio-26 con una vida media de 7,17 × 10 (717 mil años) y el potasio-40 con una vida media de 1.250 × 10 (1,25 mil millones) años, isótopos producidos principalmente en supernovas, eran mucho más comunes.
Los períodos de tiempo entre tales eventos ambientales devastadores dan ventanas de tiempo para el posible origen de la vida en los primeros entornos. Si el entorno hidrotermal marino profundo era el sitio para el origen de la vida, entonces la abiogénesis podría haber ocurrido tan pronto como 4.0 a 4.2 Ga. Si el sitio estaba en la superficie de la Tierra, la abiogénesis solo podría haber ocurrido entre 3.7 y 4.0 Ga.
En 2016, se identificó un conjunto de 355 genes probablemente presentes en el Último Ancestro Común Universal (LUCA) de todos los organismos que viven en la Tierra. Se secuenciaron un total de 6,1 millones de genes codificantes de proteínas procariotas de varios árboles filogenéticos, identificando 355 agrupaciones de proteínas de entre 286,514 agrupaciones de proteínas que probablemente eran comunes a LUCA. Los resultados "muestran a LUCA como anaeróbico, fijación de CO 2 , dependiente de H 2 con una vía Wood-Ljungdahl, N 2-fijación y termófila La bioquímica de LUCA estaba repleta de clústeres de FeS y mecanismos de reacción radicales. Sus cofactores revelan la dependencia de metales de transición, flavinas, S-adenosil metionina, coenzima A, ferredoxina, molibdopterina, corrinas y selenio. Su código genético requiere modificaciones de nucleósidos y metilaciones dependientes de S-adenosilmetionina. "Los resultados muestran clostridios metanogénicos como clado basal en las 355 filogenias examinadas, y sugieren que LUCA habitó un entorno de ventilación hidrotermal anaeróbica en un ambiente geoquímicamente activo rico en H 2 , CO 2 y hierro. MD Brazier ha demostrado que los pequeños fósiles descubiertos provienen de un mundo venenoso caliente de los gases tóxicos metano, amoníaco, dióxido de carbono y sulfuro de hidrógeno. Un análisis del árbol triple de la vida convencional muestra que las bacterias termófilas e hipertermófilas y las arqueas son las más cercanas a la raíz, lo que sugiere que la vida puede haber evolucionado en un ambiente caluroso.
Historia conceptual
Generación espontánea
La creencia en la generación espontánea de ciertas formas de vida a partir de la materia no vivida se remonta a Aristóteles y la filosofía griega antigua y continuó teniendo apoyo en la erudición occidental hasta el siglo XIX. Esta creencia se combinó con una creencia en la heterogenesis, es decir, que una forma de vida se deriva de una forma diferente (por ejemplo, las abejas de las flores). Las nociones clásicas de generación espontánea sostuvieron que ciertos organismos vivos complejos son generados por sustancias orgánicas en descomposición. Según Aristóteles, era una verdad fácilmente observable que los áfidos surgen del rocío que cae sobre las plantas, las moscas de la materia pútrida, los ratones del heno sucio, los cocodrilos de los troncos podridos en el fondo de los cuerpos de agua, y así sucesivamente. En el siglo XVII, la gente comenzó a cuestionar tales suposiciones. En 1646, Sir Thomas Browne publicó su Pseudodoxia Epidemica ( Investigacionessubtituladas sobre muchos principios recibidos, y comúnmente supuestas verdades ), que era un ataque a creencias falsas y "errores vulgares". Su contemporáneo, Alexander Ross, lo refutó erróneamente, afirmando: "cuestionar esta [generación espontánea], es cuestionar la razón, el sentido y la experiencia: si él duda de esto, déjelo ir a Egipto , y allí va a encontrar los campos. pululando con ratones engendrados del barro de Nylus , a la gran calamidad de los Habitantes ".
En 1665, Robert Hooke publicó los primeros dibujos de un microorganismo. Hooke fue seguido en 1676 por Antonie van Leeuwenhoek, quien dibujó y describió microorganismos que ahora se cree que han sido protozoos y bacterias. Muchos sintieron que la existencia de microorganismos era evidencia en apoyo de la generación espontánea, ya que los microorganismos parecían demasiado simplistas para la reproducción sexual, y la reproducción asexual a través de la división celular aún no se había observado. Van Leeuwenhoek discrepó con las ideas comunes en el momento en que las pulgas y los piojos podrían resultar espontáneamente de la putrefacción, y que las ranas también podrían surgir de la baba. Utilizando una amplia gama de experimentos que van desde la incubación de carne sellada y abierta y el estudio detallado de la reproducción de insectos, en la década de 1680 se convenció de que la generación espontánea era incorrecta.
La primera evidencia experimental contra la generación espontánea llegó en 1668 cuando Francesco Redi demostró que no aparecían gusanos en la carne cuando se impedía que las moscas pusieran huevos. Poco a poco se demostró que, al menos en el caso de todos los organismos superiores y fácilmente visibles, el sentimiento anterior con respecto a la generación espontánea era falso. La alternativa parecía ser la biogénesis: que todo ser viviente provenía de un ser viviente preexistente ( omne vivum ex ovo , latín para "cada cosa viviente de un huevo").
En 1768, Lazzaro Spallanzani demostró que los microbios estaban presentes en el aire y podían morir por ebullición. En 1861, Louis Pasteur realizó una serie de experimentos que demostraron que organismos como bacterias y hongos no aparecen espontáneamente en medios estériles ricos en nutrientes, sino que solo podían aparecer por invasión externa.
La creencia de que el auto ordenamiento por generación espontánea era imposible suplicaba una alternativa. A mediados del siglo XIX, la teoría de la biogénesis había acumulado tanto apoyo probatorio, debido al trabajo de Pasteur y otros, que la teoría alternativa de la generación espontánea había sido refutada efectivamente. John Desmond Bernal, un pionero en la cristalografía de rayos X, sugirió que las teorías anteriores, como la generación espontánea, se basaban en una explicación de que la vida se creaba continuamente como resultado de eventos fortuitos.
Etimología
El término biogénesis suele atribuirse a Henry Charlton Bastian oa Thomas Henry Huxley. Bastian usó el término alrededor de 1869 en un intercambio inédito con John Tyndall para significar "originación de vida o comienzo". En 1870, Huxley, como nuevo presidente de la Asociación Británica para el Avance de la Ciencia, pronunció un discurso titulado Biogénesis y Abiogénesis . En él, introdujo el término biogénesis (con un significado opuesto al de Bastian), así como abiogénesis :
- Y así la hipótesis de que la materia viva siempre surge por la acción de la materia viva preexistente, tomó forma definida; y tenía, de ahora en adelante, un derecho a ser considerado y una reclamación a ser refutada, en cada caso particular, antes de que la producción de materia viva de cualquier otra manera pudiera ser admitida por razonadores cuidadosos. Me será necesario referirme a esta hipótesis con tanta frecuencia, que, para salvar la circunlocución, la llamaré hipótesis de la biogénesis ; y denominaré la doctrina contraria, que la materia viva puede ser producida por la materia no viviente, la hipótesis de la abiogénesis .
Posteriormente, en el prefacio al libro de Bastian de 1871, Los modos de origen de los organismos más bajos , Bastian se refirió a la posible confusión con el uso de Huxley y renunció explícitamente a su propio significado:
- Una palabra de explicación parece necesaria con respecto a la introducción del nuevo término Arquebiosis . Originalmente, en escritos inéditos, adopté la palabra Biogénesis para expresar el mismo significado, a saber, el origen de la vida o el comienzo. Pero, mientras tanto, la palabra Biogénesis ha sido utilizada, de forma bastante independiente, por un biólogo distinguido [Huxley], que deseaba que tuviera un significado totalmente diferente. También introdujo la palabra Abiogénesis. Sin embargo, me han informado, con la mejor autoridad, que ninguna de estas palabras puede, con respecto al lenguaje del que derivan, suponer que tienen los significados que últimamente se les han asignado públicamente. Deseando evitar toda confusión innecesaria, por lo tanto, renuncié al uso de la palabra Biogénesis y, por el motivo que acabo de exponer, incapaz de adoptar el otro término, me vi obligado a introducir una nueva palabra para designar el proceso por el cual se supone que la materia viviente se forma, independientemente de la materia viva preexistente.
Louis Pasteur y Charles Darwin

Louis Pasteur comentó, sobre un hallazgo suyo en 1864 que consideró definitivo: "Nunca se recuperará la doctrina de la generación espontánea del golpe mortal golpeado por este simple experimento". Una alternativa era que los orígenes de la vida en la Tierra provenían de algún otro lugar en el universo. Periódicamente resucitado (ver Panspermia, arriba), Bernal dijo que este enfoque "es equivalente en última instancia a afirmar el funcionamiento de entidades metafísicas y espirituales ... se convierte en el argumento de la creación por diseño de un creador o demiurgo". Tal teoría, dijo Bernal, no era científica. Una teoría popular al mismo tiempo era que la vida era el resultado de una "fuerza de vida" interior, que a fines del siglo XIX fue defendida por Henri Bergson.
La idea de la evolución por selección natural propuesta por Charles Darwin puso fin a estas teologías metafísicas. En una carta a Joseph Dalton Hooker el 1 de febrero de 1871, Darwin discutió la sugerencia de que la chispa original de la vida pudo haber comenzado en un "pequeño estanque cálido, con todo tipo de sales amoniacales y fosfóricas, luz, calor, electricidad, etc." presente, que un compuesto de proteina se formó químicamente listo para someterse a cambios aún más complejos ". Continuó explicando que "en la actualidad, ese asunto sería devorado o absorbido instantáneamente, lo que no hubiera sido el caso antes de que se formaran las criaturas vivientes". Le escribió a Hooker en 1863 declarando que "es una mera basura, pensando en el presente el origen de la vida, uno podría pensar en el origen de la materia". , se había referido a que la vida había sido "creada", por lo cual "realmente quería decir 'apareció' por algún proceso totalmente desconocido", pero pronto se arrepintió de usar el término "creación" del Antiguo Testamento.
Hipótesis de "sopa primordial"

Ninguna nueva investigación notable o hipótesis sobre el tema apareció hasta 1924, cuando Alexander Oparin razonó que el oxígeno atmosférico impide la síntesis de ciertos compuestos orgánicos que son bloques de construcción necesarios para la evolución de la vida. En su libro El origen de la vidaOparin propuso que la "generación espontánea de la vida" atacada por Louis Pasteur de hecho ocurrió una vez, pero ahora era imposible porque las condiciones encontradas en la Tierra primitiva habían cambiado, y los organismos preexistentes consumirían inmediatamente cualquier organismo generado espontáneamente. Oparin argumentó que una "sopa primigenia" de moléculas orgánicas podría crearse en una atmósfera sin oxígeno a través de la acción de la luz solar. Estos se combinarían de formas cada vez más complejas hasta que formaran coacervadas. Estas gotitas "crecerían" por fusión con otras gotitas y se "reproducirían" a través de la fisión en gotas hijas, y así tienen un metabolismo primitivo en el que sobreviven factores que promueven la "integridad celular" y aquellos que no se extinguen.
Robert Shapiro ha resumido la teoría de la "sopa primordial" de Oparin y JBS Haldane en su "forma madura" de la siguiente manera:
- La Tierra primitiva tenía una atmósfera químicamente reductora.
- Esta atmósfera, expuesta a la energía en diversas formas, produce compuestos orgánicos simples ("monómeros").
- Estos compuestos se acumulan en una "sopa" que puede haberse concentrado en varios lugares (costas, respiraderos oceánicos, etc.).
- Mediante una transformación adicional, se desarrollaron en la sopa polímeros orgánicos más complejos, y en última instancia, la vida.
Aproximadamente en esta época, Haldane sugirió que los océanos prebióticos de la Tierra (bastante diferentes de sus homólogos modernos) habrían formado una "sopa caliente diluida" en la que los compuestos orgánicos podrían haberse formado. Bernal llamó a esta idea biopoiesis o biopoesis , el proceso de la materia viva que evoluciona a partir de moléculas autorreplicantes pero no vivas, y propuso que la biopoyesis pasa por una serie de etapas intermedias.
Una de las piezas más importantes de apoyo experimental para la teoría de la "sopa" llegó en 1952. Stanley L. Miller y Harold C. Urey realizaron un experimento que demostró cómo las moléculas orgánicas podrían haberse formado espontáneamente a partir de precursores inorgánicos en condiciones como las postuladas por el Hipótesis Oparin-Haldane. El ahora famoso experimento de Miller-Urey usó una mezcla altamente reductora de gases: metano, amoníaco e hidrógeno, así como también vapor de agua, para formar simples monómeros orgánicos como los aminoácidos. La mezcla de gases se cicló a través de un aparato que entregó chispas eléctricas a la mezcla. Después de una semana, se descubrió que aproximadamente del 10% al 15% del carbono en el sistema estaba entonces en forma de una mezcla racémica de compuestos orgánicos, incluidos los aminoácidos, que son los componentes básicos de las proteínas.
Bernal demostró que, en base a este y al posterior trabajo, no hay dificultad en principio para formar la mayoría de las moléculas que reconocemos como las moléculas necesarias para la vida a partir de sus precursores inorgánicos. La hipótesis subyacente sostenida por Oparin, Haldane, Bernal, Miller y Urey, por ejemplo, era que las condiciones múltiples en la Tierra primigenia favorecían las reacciones químicas que sintetizaban el mismo conjunto de compuestos orgánicos complejos a partir de tales precursores simples. Un nuevo análisis de 2011 de los viales guardados que contienen los extractos originales que resultaron de los experimentos de Miller y Urey, utilizando tecnología y equipos analíticos actuales y más avanzados, ha descubierto más productos bioquímicos que los descubiertos originalmente en la década de 1950. Uno de los hallazgos más importantes fue de 23 aminoácidos, mucho más que los cinco encontrados originalmente. Sin embargo, Bernal dijo que "
Estudios más recientes, en octubre de 2017, respaldan la noción de que la vida pudo haber comenzado justo después de que la Tierra se formara como moléculas de ARN que emergían de "pequeños estanques cálidos".
Microesferas proteinoides
Al tratar de descubrir las etapas intermedias de la abiogénesis mencionadas por Bernal, Sidney W. Fox en los años 1950 y 1960 estudió la formación espontánea de estructuras peptídicas (pequeñas cadenas de aminoácidos) en condiciones que posiblemente hubieran existido al principio de la historia de la Tierra. En uno de sus experimentos, permitió que los aminoácidos se secaran como encharcados en un lugar cálido y seco en condiciones prebióticas. Descubrió que, a medida que se secaban, los aminoácidos formaban moléculas polipeptídicas submicroscópicas largas, a menudo reticuladas, parecidas a hilos, ahora denominadas "microesferas proteinoides".
En otro experimento para establecer las condiciones adecuadas para que se forme la vida, Fox recolectó material volcánico de un cono de ceniza en Hawai. Descubrió que la temperatura era superior a 100 ° C (212 ° F) a solo 4 pulgadas (100 mm) por debajo de la superficie del cono de ceniza, y sugirió que este podría haber sido el entorno en el que se creó la vida: las moléculas podrían haberse formado y luego se lavó a través de la ceniza volcánica suelta en el mar. Colocó grumos de lava sobre los aminoácidos derivados del metano, el amoníaco y el agua, esterilizó todos los materiales y horneó la lava sobre los aminoácidos durante unas horas en un horno de vidrio. Una sustancia marrón y pegajosa se formó sobre la superficie, y cuando la lava se empapó en agua esterilizada, un líquido espeso y marrón se filtró. Los aminoácidos se habían combinado para formar proteinoides, y los proteinoides se habían combinado para formar pequeños glóbulos que Fox llamó "microesferas". Sus proteinoides no eran células, aunque formaban grupos y cadenas que recordaban a las cianobacterias, pero no contenían ácidos nucleicos funcionales ni ninguna información codificada. Basado en tales experimentos, Colin S. Pittendrigh declaró en diciembre de 1967 que "los laboratorios crearán una célula viva dentro de diez años", una observación que refleja la ingenuidad contemporánea típica sobre la complejidad de las estructuras celulares.
Modelos actuales
No existe un modelo único y generalmente aceptado para el origen de la vida. Los científicos han propuesto varias hipótesis plausibles, que comparten algunos elementos comunes. Aunque difieren en los detalles, estas hipótesis se basan en el marco establecido por Alexander Oparin (en 1924) y por JBS Haldane (en 1925), quien postuló la teoría de la evolución molecular o química de la vida. Según ellos, las primeras moléculas que constituyen las primeras células "se sintetizaron en condiciones naturales mediante un lento proceso de evolución molecular, y estas moléculas se organizaron en el primer sistema molecular con propiedades con orden biológico". Oparin y Haldane sugirieron que la atmósfera de la Tierra primitiva pudo haber sido químicamente de naturaleza reductora, compuesta principalmente de metano (CH 4 ), amoníaco (NH 3).), agua (H 2 O), sulfuro de hidrógeno (H 2 S), dióxido de carbono (CO 2 ) o monóxido de carbono (CO) y fosfato (PO 4), con oxígeno molecular (O 2 ) y ozono (O 3 ) ya sea raro o ausente. Según modelos posteriores, la atmósfera en el período tardío de los Andes consistió principalmente en nitrógeno (N 2 ) y dióxido de carbono, con cantidades más pequeñas de monóxido de carbono, hidrógeno (H 2) y compuestos de azufre; aunque carecía de oxígeno molecular y ozono, no era tan químicamente reductor como supusieron Oparin y Haldane. En la atmósfera propuesta por Oparin y Haldane, la actividad eléctrica puede producir ciertas moléculas pequeñas (monómeros) de la vida, como los aminoácidos. El experimento Miller-Urey reportado en 1953 lo demostró.
Bernal acuñó el término biopoiesis en 1949 para referirse al origen de la vida. En 1967, sugirió que ocurrió en tres "etapas":
- el origen de los monómeros biológicos
- el origen de los polímeros biológicos
- la evolución de las moléculas a las células
Bernal sugirió que la evolución comenzó entre las etapas 1 y 2. Bernal consideraba que la tercera etapa -descubrir los métodos mediante los cuales las reacciones biológicas se incorporaban detrás de los límites de una célula- era la más difícil. El trabajo moderno sobre la forma en que las membranas celulares se autoensamblan, y el trabajo sobre microporos en varios sustratos, puede ser una solución intermedia para el desarrollo de células independientes de vida libre.
A partir de la evolución química vino la iniciación de la evolución biológica, que condujo a las primeras células. Nadie ha sintetizado aún una "protocélula" utilizando componentes simples con las propiedades de vida necesarias (el llamado "enfoque ascendente"). Sin tal prueba de principio, las explicaciones han tendido a centrarse en la quimiosíntesis. Sin embargo, algunos investigadores trabajan en este campo, especialmente Steen Rasmussen y Jack W. Szostak. Otros han argumentado que un "enfoque de arriba hacia abajo" es más factible. Uno de estos enfoques, exitosamente intentado por Craig Venter y otros en J. Craig Venter Institute, involucra la ingeniería de las células procariotas existentes con un número cada vez menor de genes, tratando de discernir en qué punto se alcanzan los requisitos más mínimos para la vida.
La estrategia de la NASA sobre abiogénesis establece que es necesario identificar interacciones, estructuras y funciones intermedias, fuentes de energía y factores ambientales que contribuyeron a la diversidad, selección y replicación de sistemas macromoleculares evolutivos. El énfasis debe continuar para mapear el panorama químico de los potenciales polímeros de información primordiales. El advenimiento de polímeros que podrían replicar, almacenar información genética y exhibir propiedades sujetas a selección probablemente fue un paso crítico en la aparición de la evolución química prebiótica.
Origen químico de moléculas orgánicas
Los elementos, a excepción del hidrógeno y el helio, derivan en última instancia de la nucleosíntesis estelar. El 12 de octubre de 2016, los astrónomos informaron que los ingredientes químicos básicos de la vida, la molécula de carbono-hidrógeno (CH o radical metilidino), el ion positivo de carbono-hidrógeno (CH +) y el ion de carbono (C +), son el resultado de luz ultravioleta de estrellas, en lugar de otras formas de radiación de supernovas y estrellas jóvenes, como se pensó anteriormente. Las moléculas complejas, incluidas las moléculas orgánicas, se forman naturalmente tanto en el espacio como en los planetas. Hay dos posibles fuentes de moléculas orgánicas en la Tierra primitiva:
- Orígenes terrestres: síntesis de moléculas orgánicas impulsada por choques de impacto u otras fuentes de energía (como luz UV, acoplamiento redox o descargas eléctricas, por ejemplo, los experimentos de Miller)
- Orígenes extraterrestres: formación de moléculas orgánicas en las nubes de polvo interestelar, que caen sobre los planetas. (Ver pseudopanspermia)
Con base en estudios recientes de modelos computarizados, las complejas moléculas orgánicas necesarias para la vida se pueden haber formado en el disco protoplanetario de los granos de polvo que rodean al Sol antes de la formación de la Tierra. Según los estudios por computadora, este mismo proceso también puede ocurrir alrededor de otras estrellas que adquieren planetas. (También vea moléculas orgánicas extraterrestres).
Las estimaciones de la producción de compuestos orgánicos a partir de estas fuentes sugieren que el Bombardeo Pesado Tardío antes de 3,5 Ga en la atmósfera primitiva proporcionó cantidades de compuestos orgánicos comparables a las producidas por fuentes terrestres.
Se ha estimado que el Bombardeo Pesado Tardío también puede haber esterilizado efectivamente la superficie de la Tierra a una profundidad de decenas de metros. Si la vida evolucionara más profundamente que esto, también se habría protegido de los altos niveles iniciales de radiación ultravioleta del estadio T Tauri de la evolución del Sol. Las simulaciones de la corteza oceánica calentada geotérmicamente producen muchos más elementos orgánicos que los encontrados en los experimentos de Miller-Urey (ver abajo). En las profundas fuentes hidrotermales, Everett Shock ha descubierto que "existe un enorme impulso termodinámico para formar compuestos orgánicos, como el agua de mar y los fluidos hidrotermales, que están lejos del equilibrio, se mezclan y se mueven hacia un estado más estable". Shock ha descubierto que la energía disponible se maximiza a alrededor de 100 - 150 grados Celsius,
La acumulación y concentración de moléculas orgánicas en una superficie planetaria también se considera un paso inicial esencial para el origen de la vida. Identificar y comprender los mecanismos que condujeron a la producción de moléculas prebióticas en diversos ambientes es fundamental para establecer el inventario de ingredientes a partir de los cuales se originó la vida en la Tierra, suponiendo que la producción abiótica de moléculas influyó finalmente en la selección de moléculas de las que surgió la vida.
Síntesis química
Si bien las características de la autoorganización y la autorreplicación a menudo se consideran el sello distintivo de los sistemas vivos, existen muchas instancias de moléculas abióticas que exhiben tales características en condiciones adecuadas. Stan Palasek sugirió basado en un modelo teórico que el autoensamblaje de moléculas de ácido ribonucleico (ARN) puede ocurrir espontáneamente debido a factores físicos en los respiraderos hidrotermales. El autoensamblaje del virus dentro de las células anfitrionas tiene implicaciones para el estudio del origen de la vida, ya que da mayor credibilidad a la hipótesis de que la vida podría haber comenzado como moléculas orgánicas autoensambladas.
Múltiples fuentes de energía estaban disponibles para reacciones químicas en la Tierra primitiva. Por ejemplo, el calor (como los procesos geotérmicos) es una fuente de energía estándar para la química. Otros ejemplos incluyen luz solar y descargas eléctricas (rayos), entre otros. Las simulaciones por computadora también sugieren que la cavitación en los depósitos de agua primordiales, tales como la ruptura de las olas del mar, los arroyos y los océanos, puede potencialmente conducir a la síntesis de compuestos biogénicos. Las reacciones desfavorables también pueden ser impulsadas por reacciones altamente favorables, como en el caso de la química de hierro y azufre. Por ejemplo, esto fue probablemente importante para la fijación de carbono (la conversión de carbono de su forma inorgánica a una orgánica). La fijación de carbono a través de la química de hierro y azufre es altamente favorable y ocurre a pH neutro y 100 ° C (212 ° F). Superficies de hierro y azufre
Ya en la década de 1860, los experimentos han demostrado que las moléculas biológicamente relevantes se pueden producir a partir de la interacción de fuentes simples de carbono con abundantes catalizadores inorgánicos. En particular, los experimentos de Butlerov (la reacción formosa) mostraron que las tetrosas, las pentosas y las hexosas se producen cuando el formaldehído se calienta en condiciones básicas con iones metálicos divalentes como el calcio. La reacción fue analizada y posteriormente se propuso que sería autocatalítica por Breslow en 1959. Experimentos similares (ver a continuación) demuestran que las nucleobases como la guanina y la adenina podrían sintetizarse a partir de fuentes simples de carbono y nitrógeno como el cianuro de hidrógeno y el amoníaco.
La formamida produce los cuatro ribonucleótidos y otras moléculas biológicas cuando se calienta en presencia de varios minerales terrestres. La formamida es ubicua en el Universo, producida por la reacción del agua y el cianuro de hidrógeno (HCN). Tiene varias ventajas como precursor biótico, incluida la capacidad de concentrarse fácilmente a través de la evaporación del agua. Aunque el HCN es venenoso, solo afecta a los organismos aeróbicos (eucariotas y bacterias aeróbicas), que aún no existían. También puede desempeñar funciones en otros procesos químicos, como la síntesis del aminoácido glicina.
En 1961, se demostró que la adenina de ácido nucleico purina se puede formar calentando soluciones acuosas de cianuro de amonio. También se informaron otras vías para sintetizar bases a partir de materiales inorgánicos. Leslie E. Orgel y sus colegas han demostrado que las temperaturas de congelación son ventajosas para la síntesis de purinas, debido al efecto concentrador de precursores clave como el cianuro de hidrógeno. La investigación de Stanley L. Miller y sus colegas sugirieron que, si bien la adenina y la guanina requieren condiciones de congelación para la síntesis, la citosina y el uracilo pueden requerir temperaturas de ebullición. La investigación realizada por el grupo Miller señala la formación de siete aminoácidos diferentes y 11 tipos de nucleobases en hielo cuando el amoníaco y el cianuro se dejaron en un congelador entre 1972 y 1997. Otro trabajo demostró la formación de s-triazinas (nucleobases alternativas), pirimidinas (incluyendo citosina y uracilo) y adenina de soluciones de urea bajo ciclos de congelación-descongelación bajo una atmósfera reductiva (con descargas de chispas como fuente de energía). La explicación dada para la velocidad inusual de estas reacciones a una temperatura tan baja es la congelación eutéctica. A medida que se forma un cristal de hielo, se mantiene puro: solo las moléculas de agua se unen al cristal en crecimiento, mientras que las impurezas como la sal o el cianuro quedan excluidas. Estas impurezas se llenan en bolsas microscópicas de líquido dentro del hielo, y esta aglomeración hace que las moléculas colisionen más a menudo. La exploración mecánica utilizando métodos químicos cuánticos proporciona una comprensión más detallada de algunos de los procesos químicos implicados en la evolución química, y una respuesta parcial a la cuestión fundamental de la biogénesis molecular. y adenina a partir de soluciones de urea sometidas a ciclos de congelación-descongelación bajo una atmósfera reductiva (con descargas de chispas como fuente de energía). La explicación dada para la velocidad inusual de estas reacciones a una temperatura tan baja es la congelación eutéctica. A medida que se forma un cristal de hielo, se mantiene puro: solo las moléculas de agua se unen al cristal en crecimiento, mientras que las impurezas como la sal o el cianuro quedan excluidas. Estas impurezas se llenan en bolsas microscópicas de líquido dentro del hielo, y esta aglomeración hace que las moléculas colisionen más a menudo. La exploración mecánica utilizando métodos químicos cuánticos proporciona una comprensión más detallada de algunos de los procesos químicos implicados en la evolución química, y una respuesta parcial a la cuestión fundamental de la biogénesis molecular. y adenina a partir de soluciones de urea sometidas a ciclos de congelación-descongelación bajo una atmósfera reductiva (con descargas de chispas como fuente de energía). La explicación dada para la velocidad inusual de estas reacciones a una temperatura tan baja es la congelación eutéctica. A medida que se forma un cristal de hielo, se mantiene puro: solo las moléculas de agua se unen al cristal en crecimiento, mientras que las impurezas como la sal o el cianuro quedan excluidas. Estas impurezas se llenan en bolsas microscópicas de líquido dentro del hielo, y esta aglomeración hace que las moléculas colisionen más a menudo. La exploración mecánica utilizando métodos químicos cuánticos proporciona una comprensión más detallada de algunos de los procesos químicos implicados en la evolución química, y una respuesta parcial a la cuestión fundamental de la biogénesis molecular.
En el momento del experimento Miller-Urey, el consenso científico era que la Tierra primitiva tenía una atmósfera reductora con compuestos relativamente ricos en hidrógeno y pobres en oxígeno (p. Ej., CH 4 y NH 3 en comparación con CO 2 y dióxido de nitrógeno (NO 2))). Sin embargo, el consenso científico actual describe la atmósfera primitiva como débilmente reductora o neutral (véase también la catástrofe de oxígeno). Tal atmósfera disminuiría tanto la cantidad como la variedad de aminoácidos que podrían producirse, aunque los estudios que incluyen minerales de hierro y carbonato (pensamiento presente en los océanos tempranos) en las condiciones experimentales han producido una serie diversa de aminoácidos. Otra investigación científica se ha centrado en otros dos posibles entornos reductores: el espacio exterior y los respiraderos térmicos de aguas profundas.
La formación espontánea de polímeros complejos a partir de monómeros generados abióticamente en las condiciones planteadas por la teoría de la "sopa" no es en absoluto un proceso directo. Además de los monómeros orgánicos básicos necesarios, los compuestos que habrían prohibido la formación de polímeros también se formaron en alta concentración durante los experimentos de Miller-Urey y Joan Oró. El experimento Miller-Urey, por ejemplo, produce muchas sustancias que reaccionarían con los aminoácidos o terminarían su acoplamiento en cadenas peptídicas.
Un proyecto de investigación completado en marzo de 2015 por John D. Sutherland y otros descubrió que una red de reacciones que comenzaba con cianuro de hidrógeno y sulfuro de hidrógeno, en corrientes de agua irradiadas con luz UV, podía producir componentes químicos de proteínas y lípidos, así como los de ARN, aunque no producen una amplia gama de otros compuestos. Los investigadores usaron el término "ciano sulfúrico" para describir esta red de reacciones.
Autocatálisis
Los autocatalizadores son sustancias que catalizan la producción de sí mismos y, por lo tanto, son "replicadores moleculares". Los sistemas químicos autorreplicantes más simples son autocatalíticos, y típicamente contienen tres componentes: una molécula de producto y dos moléculas precursoras. La molécula del producto une las moléculas precursoras, que a su vez producen más moléculas producto de más moléculas precursoras. La molécula producto cataliza la reacción proporcionando una plantilla complementaria que se une a los precursores, uniéndolos. Tales sistemas se han demostrado tanto en macromoléculas biológicas como en moléculas orgánicas pequeñas. También se han observado sistemas que no proceden por mecanismos de plantilla, como la autorreproducción de micelas y vesículas.
Se ha propuesto que la vida surgió inicialmente como redes químicas autocatalíticas. El etólogo británico Richard Dawkins escribió sobre la autocatálisis como una posible explicación del origen de la vida en su libro de 2004 The Ancestor's Tale . En su libro, Dawkins cita experimentos realizados por Julius Rebek Jr. y sus colegas en los que combinaron amino adenosina y pentafluorofenil ésteres con el autocatalizador amino adenosina triácido éster (AATE). Un producto era una variante de AATE, que catalizaba la síntesis de ellos mismos. Este experimento demostró la posibilidad de que los autocatalizadores pudieran exhibir competencia dentro de una población de entidades con herencia, lo que podría interpretarse como una forma rudimentaria de selección natural.
A principios de la década de 1970, Manfred Eigen y Peter Schuster examinaron las etapas transitorias entre el caos molecular y un hiperciclo autorreplicante en una sopa prebiótica. En un hiperciclo, el sistema de almacenamiento de información (posiblemente ARN) produce una enzima, que cataliza la formación de otro sistema de información, en secuencia hasta que el producto de los últimos ayuda en la formación del primer sistema de información. Los hiperciclos tratados matemáticamente podrían crear cuasiespecies, que a través de la selección natural entraron en una forma de evolución darwiniana. Un impulso a la teoría del hiperciclo fue el descubrimiento de ribozimas capaces de catalizar sus propias reacciones químicas. La teoría del hiperciclo requiere la existencia de bioquímicos complejos, como nucleótidos, que no se forman en las condiciones propuestas por el experimento de Miller-Urey.
Geoffrey W. Hoffmann ha demostrado que una maquinaria de traducción temprana propensa a errores puede ser estable frente a una catástrofe de error del tipo que se había previsto como problemático para el origen de la vida, y era conocida como "la paradoja de Orgel".
Hoffmann también ha argumentado que un evento de nucleación complejo como el origen de la vida que involucra tanto polipéptidos como ácido nucleico es compatible con el tiempo y el espacio disponible en los océanos primitivos de la Tierra Hoffmann sugiere que la ceniza volcánica puede proporcionar las muchas formas aleatorias necesarias en el complejo postulado evento de nucleación. Este aspecto de la teoría puede probarse experimentalmente.
Homoquiralidad
La homoquiralidad se refiere a la uniformidad geométrica de algunos materiales compuestos de unidades quirales. Chiral se refiere a formas 3D no superponibles que son imágenes especulares entre sí, al igual que las manos izquierda y derecha. Los organismos vivos usan moléculas que tienen la misma quiralidad ("lateralidad"): casi sin excepciones, los aminoácidos son zurdos, mientras que los nucleótidos y azúcares son diestros. Las moléculas quirales se pueden sintetizar, pero en ausencia de una fuente quiral o un catalizador quiral, se forman en una mezcla 50/50 de ambos enantiómeros (denominada mezcla racémica). Los mecanismos conocidos para la producción de mezclas no racémicas a partir de materiales de partida racémicos incluyen: leyes físicas asimétricas, tales como la interacción electrodébil; ambientes asimétricos, como los causados por la luz polarizada circularmente, cristales de cuarzo o la rotación de la Tierra,
Una vez establecido, se seleccionará quiralidad para. Un pequeño sesgo (exceso enantiomérico) en la población se puede amplificar en uno grande por autocatálisis asimétrica, como en la reacción de Soai. En la autocatálisis asimétrica, el catalizador es una molécula quiral, lo que significa que una molécula quiral está catalizando su propia producción. Un exceso enantiomérico inicial, como el que puede ser producido por la luz polarizada, permite que el enantiómero más abundante supere al otro.
Clark ha sugerido que la homoquiralidad puede haber comenzado en el espacio exterior, ya que los estudios de los aminoácidos en el meteorito Murchison mostraron que la L-alanina es más del doble de frecuente que la forma D, y que el ácido L-glutámico era más de tres veces prevalente que su contraparte D Diversas superficies de cristales quirales también pueden actuar como sitios para la posible concentración y ensamblaje de unidades monoméricas quirales en macromoléculas. Los compuestos encontrados en los meteoritos sugieren que la quiralidad de la vida se deriva de la síntesis abiogénica, ya que los aminoácidos de los meteoritos muestran un sesgo zurdo, mientras que los azúcares muestran un sesgo predominantemente diestro, el mismo que se encuentra en los organismos vivos.
Autocierre, reproducción, duplicación y el mundo del ARN
Protoceldas

Una protocélula es una colección de lípidos autoorganizada, autoordenada y esférica propuesta como un trampolín hacia el origen de la vida. Una cuestión central en la evolución es cómo las protocélulas simples surgieron por primera vez y diferían en la contribución reproductiva a la siguiente generación que impulsa la evolución de la vida. Aunque todavía no se ha logrado una protocélula funcional en un entorno de laboratorio, hay científicos que piensan que el objetivo está bien al alcance.
Las vesículas autoensambladas son componentes esenciales de las células primitivas. La segunda ley de la termodinámica requiere que el universo se mueva en una dirección en la que la entropía aumenta, aunque la vida se distingue por su gran grado de organización. Por lo tanto, se necesita un límite para separar los procesos de la vida de la materia no viva. Los investigadores Irene A. Chen y Jack W. Szostak, entre otros, sugieren que las propiedades fisicoquímicas simples de las protocélulas elementales pueden dar lugar a comportamientos celulares esenciales, incluidas formas primitivas de competencia de reproducción diferencial y almacenamiento de energía. Tales interacciones cooperativas entre la membrana y sus contenidos encapsulados podrían simplificar en gran medida la transición de simples moléculas de replicación a células verdaderas. Además, la competencia por las moléculas de membrana favorecería a las membranas estabilizadas, sugiriendo una ventaja selectiva para la evolución de los ácidos grasos reticulados e incluso los fosfolípidos de hoy. Tal microencapsulación permitiría el metabolismo dentro de la membrana, el intercambio de moléculas pequeñas, pero la prevención del paso de sustancias grandes a través de ella. Las principales ventajas de la encapsulación incluyen la mayor solubilidad de la carga contenida dentro de la cápsula y el almacenamiento de energía en forma de un gradiente electroquímico.
Un estudio de 2012 dirigido por Armen Y. Mulkidjanian de la Universidad alemana de Osnabrück, sugiere que las piscinas interiores de vapor geotérmico condensado y refrigerado tienen las características ideales para el origen de la vida. Los científicos confirmaron en 2002 que al agregar una montmorilloniteclay a una solución de micelas de ácidos grasos (esferas de lípidos), la arcilla aceleró la tasa de formación de vesículas en 100 veces.
Otro modelo de protocélula es el Jeewanu. Sintetizado por primera vez en 1963 a partir de minerales simples y elementos orgánicos básicos expuestos a la luz solar, todavía se informa que tiene algunas capacidades metabólicas, la presencia de una membrana semipermeable, aminoácidos, fosfolípidos, carbohidratos y moléculas similares al ARN. Sin embargo, la naturaleza y las propiedades de Jeewanu aún no se han aclarado.
Las interacciones electrostáticas inducidas por péptidos hidrofóbicos cortos, positivamente cargados que contienen 7 aminoácidos de longitud o menos, pueden unir ARN a una membrana vesicular, la membrana celular básica.
Mundo de ARN

La hipótesis del mundo del ARN describe una Tierra primitiva con ARN autoreplicante y catalítico, pero sin ADN ni proteínas. En general, se acepta que la vida actual en la Tierra desciende de un mundo de ARN, aunque la vida basada en ARN puede no haber sido la primera vida en existir. Esta conclusión se extrae de muchas líneas de evidencia independientes, como las observaciones de que el ARN es fundamental para el proceso de traducción y que los ARN pequeños pueden catalizar todos los grupos químicos y las transferencias de información necesarias para la vida. La estructura del ribosoma ha sido llamada la "pistola humeante", ya que demostró que el ribosoma es una ribozima, con un núcleo central de ARN y sin cadenas laterales de aminoácidos dentro de los 18 angstroms del sitio activo donde se cataliza la formación del enlace peptídico. El concepto del mundo del ARN fue propuesto por primera vez en 1962 por Alexander Rich,
Los posibles precursores para la evolución de la síntesis proteica incluyen un mecanismo para sintetizar cofactores peptídicos cortos o formar un mecanismo para la duplicación del ARN. Es probable que el ribosoma ancestral estuviera compuesto completamente de ARN, aunque algunas funciones ya han sido asumidas por las proteínas. Las principales preguntas restantes sobre este tema incluyen identificar la fuerza selectiva para la evolución del ribosoma y determinar cómo surgió el código genético.
Eugene Koonin dijo: "A pesar del considerable esfuerzo experimental y teórico, actualmente no existen escenarios convincentes para el origen de la replicación y la traducción, los procesos clave que en conjunto conforman el núcleo de los sistemas biológicos y el aparente prerrequisito de la evolución biológica. podría ofrecer la mejor oportunidad para la resolución de este acertijo, pero hasta ahora no puede explicar adecuadamente la aparición de una replicasa de ARN eficiente o el sistema de traducción. La MWO [Ed .: "muchos mundos en uno"] versión del modelo cosmológico de la eternidad la inflación podría sugerir una salida a este enigma porque, en un multiverso infinito con un número finito de historias macroscópicas distintas (cada una repetida un número infinito de veces),incluso la aparición de sistemas altamente complejos por casualidad no es solo posible sino inevitable ".
Orígenes virales
Se han sugerido pruebas recientes de una hipótesis del "virus primero", que puede respaldar las teorías del mundo del ARN. Una de las dificultades para el estudio de los orígenes de los virus es su alta tasa de mutación; este es particularmente el caso en retrovirus de ARN como el VIH. Un estudio de 2015 comparó las estructuras de pliegues de proteínas en diferentes ramas del árbol de la vida, donde los investigadores pueden reconstruir las historias evolutivas de los pliegues y de los organismos cuyos genomas codifican para esos pliegues. Ellos argumentan que los pliegues de proteínas son mejores marcadores de eventos antiguos ya que sus estructuras tridimensionales pueden mantenerse incluso cuando las secuencias que codifican para esos comiencen a cambiar. Por lo tanto, el repertorio de proteínas virales conserva rastros de la historia evolutiva antigua que se puede recuperar utilizando enfoques bioinformáticos avanzados.
Síntesis y replicación de ARN
Se han presentado varias hipótesis de formación de ARN. A partir de 1994, hubo dificultades en la explicación de la síntesis abiótica de los nucleótidos citosina y uracilo. La investigación posterior ha mostrado posibles rutas de síntesis; por ejemplo, la formamida produce los cuatro ribonucleótidos y otras moléculas biológicas cuando se calientan en presencia de diversos minerales terrestres. Las membranas celulares tempranas se podrían haber formado espontáneamente a partir de proteinoides, que son moléculas parecidas a proteínas producidas cuando las soluciones de aminoácidos se calientan en la concentración correcta de solución acuosa. Se ve que estos forman microesferas que se observa que se comportan de forma similar a los compartimentos encerrados en la membrana.
Los factores que respaldan un papel importante del ARN en la vida temprana incluyen su capacidad para actuar tanto para almacenar información como para catalizar reacciones químicas (como una ribozima); sus muchos papeles importantes como intermedio en la expresión y el mantenimiento de la información genética (en forma de ADN) en organismos modernos; y la facilidad de la síntesis química de al menos los componentes de la molécula de ARN en las condiciones que se aproximaron a la Tierra primitiva. Se han sintetizado moléculas de ARN relativamente cortas, capaces de replicarse. Tal RNA de replicasa, que funciona tanto como código como catalizador proporciona su propia plantilla sobre la cual puede ocurrir la copia. Jack W. Szostak ha demostrado que ciertos ARN catalíticos pueden unirse a secuencias de ARN más pequeñas juntas, creando el potencial de autorreplicación. Si estas condiciones estuvieran presentes, La selección natural darwiniana favorecería la proliferación de tales conjuntos autocatalíticos, a los cuales se podrían agregar otras funcionalidades. Se han identificado tales sistemas autocatalíticos de ARN capaces de replicación autosostenida. Los sistemas de replicación de ARN, que incluyen dos ribozimas que catalizan mutuamente la síntesis, mostraron un tiempo de duplicación del producto de aproximadamente una hora, y estaban sujetos a selección natural en las condiciones que existían en el experimento. En los experimentos de competencia evolutiva, esto condujo a la aparición de nuevos sistemas que se replicaron de manera más eficiente. Esta fue la primera demostración de adaptación evolutiva que se produce en un sistema genético molecular. Se han identificado tales sistemas autocatalíticos de ARN capaces de replicación autosostenida. Los sistemas de replicación de ARN, que incluyen dos ribozimas que catalizan mutuamente la síntesis, mostraron un tiempo de duplicación del producto de aproximadamente una hora, y estaban sujetos a selección natural en las condiciones que existían en el experimento. En los experimentos de competencia evolutiva, esto condujo a la aparición de nuevos sistemas que se replicaron de manera más eficiente. Esta fue la primera demostración de adaptación evolutiva que se produce en un sistema genético molecular. Se han identificado tales sistemas autocatalíticos de ARN capaces de replicación autosostenida. Los sistemas de replicación de ARN, que incluyen dos ribozimas que catalizan mutuamente la síntesis, mostraron un tiempo de duplicación del producto de aproximadamente una hora, y estaban sujetos a selección natural en las condiciones que existían en el experimento. En los experimentos de competencia evolutiva, esto condujo a la aparición de nuevos sistemas que se replicaron de manera más eficiente. Esta fue la primera demostración de adaptación evolutiva que se produce en un sistema genético molecular. En los experimentos de competencia evolutiva, esto condujo a la aparición de nuevos sistemas que se replicaron de manera más eficiente. Esta fue la primera demostración de adaptación evolutiva que se produce en un sistema genético molecular. En los experimentos de competencia evolutiva, esto condujo a la aparición de nuevos sistemas que se replicaron de manera más eficiente. Esta fue la primera demostración de adaptación evolutiva que se produce en un sistema genético molecular.
Dependiendo de la definición, la vida comenzó cuando las cadenas de ARN comenzaron a autorreplicarse, iniciando los tres mecanismos de selección darwiniana: heredabilidad, variación del tipo y producción reproductiva diferencial. La aptitud de un replicador de ARN (su tasa de aumento per cápita) probablemente dependería de sus capacidades intrínsecas de adaptación, determinadas por su secuencia de nucleótidos y la disponibilidad de recursos. Las tres capacidades adaptativas primarias pueden haber sido: (1) replicación con fidelidad moderada, lo que da lugar a heredabilidad al mismo tiempo que permite variación de tipo, (2) resistencia a la descomposición y (3) adquisición de recursos de proceso. Estas capacidades habrían funcionado por medio de las configuraciones plegadas de los replicadores de ARN resultantes de sus secuencias de nucleótidos.
Carl Zimmer ha especulado que las condiciones químicas, incluida la presencia de boro, molibdeno y oxígeno necesarios para la producción inicial de ARN, pueden haber sido mejores en Marte temprano que en la Tierra primitiva. Si es así, las moléculas adecuadas para la vida que se originan en Marte pueden haber migrado posteriormente a la Tierra a través de eyecciones de meteoritos.
Mundo previo al ARN
Es posible que un tipo diferente de ácido nucleico, como PNA, TNA o GNA, sea el primero en emerger como una molécula de autorreproducción, solo que más tarde sea reemplazado por RNA. Larralde et al. , dicen que "la síntesis prebiótica generalmente aceptada de ribosa, la reacción formosa, produce numerosos azúcares sin ninguna selectividad" y concluyen que sus "resultados sugieren que la columna vertebral del primer material genético no pudo contener ribosa u otros azúcares debido a su inestabilidad ". Se sabe que el enlace éster de la ribosa y el ácido fosfórico en el ARN es propenso a la hidrólisis.
Los ribonucleósidos de pirimidina y sus nucleótidos respectivos se han sintetizado prebióticamente mediante una secuencia de reacciones que eluden los azúcares libres, y se ensamblan de forma escalonada mediante el uso de químicas nitrogenadas u oxigenadas. Sutherland ha demostrado rutas de alto rendimiento para ribonucleótidos de citidina y uridina construidos a partir de pequeños fragmentos de carbono de 2 y 3, como glicolaldehído, gliceraldehído o gliceraldehído-3-fosfato, cianamida y cianoacetileno. Uno de los pasos de esta secuencia permite el aislamiento de la ribosa aminooxazolina enantiopura si el exceso enantiomérico de gliceraldehído es del 60% o superior. Esto puede verse como una etapa de purificación prebiótica, donde dicho compuesto cristalizó espontáneamente a partir de una mezcla de las otras pentosas aminooxazolinas. La ribosa aminooxazolina puede reaccionar con cianoacetileno de una manera suave y altamente eficiente para producir el ribonucleótido alfa citidina. La fotoanomerización con luz ultravioleta permite la inversión alrededor del centro anomérico de 1 'para proporcionar la estereoquímica beta correcta. En 2009 demostraron que los mismos bloques de construcción simples permiten el acceso, mediante la elaboración de nucleobases controladas con fosfato, a los nucleótidos de pirimidina 2 ', 3'-cíclicos directamente, que se sabe que son capaces de polimerizar en ARN. Este documento también destaca la posibilidad de foto-desinfección de los fosfatos pirimidina-2 ', 3'-cíclicos. En 2009 demostraron que los mismos bloques de construcción simples permiten el acceso, mediante la elaboración de nucleobases controladas con fosfato, a los nucleótidos de pirimidina 2 ', 3'-cíclicos directamente, que se sabe que son capaces de polimerizar en ARN. Este documento también destaca la posibilidad de foto-desinfección de los fosfatos pirimidina-2 ', 3'-cíclicos. En 2009 demostraron que los mismos bloques de construcción simples permiten el acceso, mediante la elaboración de nucleobases controladas con fosfato, a los nucleótidos de pirimidina 2 ', 3'-cíclicos directamente, que se sabe que son capaces de polimerizar en ARN. Este documento también destaca la posibilidad de foto-desinfección de los fosfatos pirimidina-2 ', 3'-cíclicos.
Origen del metabolismo biológico
Las reacciones similares al metabolismo podrían haber ocurrido naturalmente en los océanos tempranos, antes de que los primeros organismos evolucionaran. El metabolismo puede ser anterior al origen de la vida, que puede haber evolucionado a partir de las condiciones químicas en los primeros océanos. Las reconstrucciones en laboratorios muestran que algunas de estas reacciones pueden producir ARN, y algunas otras se asemejan a dos cascadas de reacción esenciales del metabolismo: la glucólisis y la vía de la pentosa fosfato, que proporcionan precursores esenciales para los ácidos nucleicos, aminoácidos y lípidos. Un estudio en la Universidad de Düsseldorf creó árboles filogénicos basados en 6 millones de genes de bacterias y arqueas, e identificó 355 familias de proteínas que probablemente estaban presentes en el LUCA. Se basaban en un metabolismo anaeróbico que fijaba el dióxido de carbono y el nitrógeno. Sugiere que el LUCA evolucionó en un ambiente rico en hidrógeno, dióxido de carbono y hierro. Los siguientes son algunos descubrimientos observados e hipótesis relacionadas.
Mundo de hierro y azufre
En la década de 1980, Günter Wächtershäuser, alentado y apoyado por Karl R. Popper, postuló su mundo de hierro y azufre, una teoría de la evolución de las vías químicas prebióticas como punto de partida en la evolución de la vida. Rastrea sistemáticamente la bioquímica de hoy en día a las reacciones primordiales que proporcionan vías alternativas a la síntesis de bloques orgánicos de compuestos gaseosos simples.
En contraste con los experimentos clásicos de Miller, que dependen de fuentes externas de energía (rayos simulados, radiación ultravioleta), los "sistemas Wächtershäuser" vienen con una fuente de energía incorporada: sulfuros de hierro (pirita de hierro) y otros minerales. La energía liberada de las reacciones redox de estos sulfuros metálicos está disponible para la síntesis de moléculas orgánicas, y tales sistemas pueden haber evolucionado en conjuntos autocatalíticos que constituyen entidades meta-replicativas metabólicamente activas anteriores a las formas de vida conocidas en la actualidad. Los experimentos con tales sulfuros en un entorno acuoso a 100 ° C produjeron un rendimiento relativamente pequeño de dipéptidos (0,4% a 12,4%) y un menor rendimiento de tripéptidos (0,10%) aunque en las mismas condiciones, los dipéptidos se descompusieron rápidamente.
Varios modelos rechazan la autorreplicación de un "gen desnudo", postulando en cambio la aparición de un metabolismo primitivo que proporciona un entorno seguro para la posterior aparición de la replicación del ARN. La centralidad del ciclo de Krebs (ciclo del ácido cítrico) para la producción de energía en organismos aeróbicos, y para atraer dióxido de carbono e iones de hidrógeno en la biosíntesis de compuestos químicos orgánicos complejos, sugiere que fue una de las primeras partes del metabolismo en evolucionar. Concordantemente, el geoquímico Michael Raquel ha propuesto que "el propósito de la vida es hidrogenar el dióxido de carbono" (como parte de un escenario "primero en el metabolismo", en lugar de "primero en genética"). El físico Jeremy England del MIT ha propuesto que la vida era inevitable a partir de consideraciones termodinámicas generales: "... cuando un grupo de átomos es impulsado por una fuente de energía externa (como el sol o combustible químico) y rodeado por un baño de calor (como el océano o la atmósfera), a menudo se reestructurará gradualmente para disipar cada vez más energía. Esto podría significar que bajo ciertas condiciones, la materia adquiere inexorablemente el atributo físico clave asociado con la vida ".
Una de las primeras encarnaciones de esta idea se presentó en 1924 con la noción de Oparin de primitivas vesículas autorreplicantes anteriores al descubrimiento de la estructura del ADN. Las variantes en los años 1980 y 1990 incluyen la teoría del mundo de hierro-azufre de Wächtershäuser y los modelos introducidos por Christian de Duve basados en la química de los tioésteres. Los argumentos más abstractos y teóricos para la verosimilitud de la aparición del metabolismo sin la presencia de genes incluyen un modelo matemático introducido por Freeman Dyson a principios de los años ochenta y la noción de conjuntos autocatalíticos colectivos de Stuart Kauffman, discutida más adelante en esa década.
Orgel resumió su análisis al afirmar: "Actualmente, no hay razón para esperar que los ciclos de varios pasos, como el ciclo del ácido cítrico reductivo, se autoorganicen en la superficie de FeS / FeS 2. o algún otro mineral. "Es posible que otro tipo de vía metabólica se usara al comienzo de la vida. Por ejemplo, en lugar del ciclo reductor de ácido cítrico, la vía" abierta "de acetil-CoA (otra de las cinco formas reconocidas) de la fijación de dióxido de carbono en la naturaleza de hoy) sería compatible con la idea de autoorganización en una superficie de sulfuro de metal. La enzima clave de esta vía, la monóxido de carbono deshidrogenasa / acetil-CoA sintasa, alberga grupos mixtos de níquel-hierro-azufre en su La reacción se centra y cataliza la formación de acetil-CoA (similar al acetil-tiol) en un solo paso. Sin embargo, existe una preocupación creciente de que los compuestos prebióticos tiolados y tioésteres sean termodinámicamente y cinéticamente desfavorables para acumularse en supuestas condiciones prebióticas (es decir, fuentes hidrotermales). )También se ha propuesto que la cisteína y la homocisteína pueden haber reaccionado con los nitrilos resultantes de la reacción de Stecker, formando fácilmente péptidos catalíticos de alcance tiolico.
Hipótesis del mundo Zn
La teoría Zn-world (zinc world) de Armen Y. Mulkidjanian es una extensión de la hipótesis de pirita de Wächtershäuser. Wächtershäuser basó su teoría de los procesos químicos iniciales que conducen a moléculas informativas (ARN, péptidos) en una malla regular de cargas eléctricas en la superficie de la pirita que puede haber facilitado la polimerización primitiva atrayendo reactivos y disponiéndolos de forma adecuada entre sí. La teoría del mundo Zn especifica y diferencia aún más. Fluidos hidrotermales ricos en H 2S interactuando con el océano primordial frío (o el "estanque cálido" de Darwin) el agua conduce a la precipitación de partículas de sulfuro de metal. Los sistemas de ventilación oceánica y otros sistemas hidrotermales tienen una estructura zonal reflejada en antiguos depósitos de sulfuros masivos volcanogénicos (VMS) de origen hidrotermal. Llegan a muchos kilómetros de diámetro y datan del Eón Arqueano. Los más abundantes son pirita (FeS 2 ), calcopirita (CuFeS 2) y esfalerita (ZnS), con adiciones de galena (PbS) y alabandita (MnS). ZnS y MnS tienen una capacidad única para almacenar energía de radiación, por ejemplo, de luz UV. Durante la ventana de tiempo relevante de los orígenes de las moléculas replicantes, la presión atmosférica primordial fue lo suficientemente alta (> 100 bar, alrededor de 100 atmósferas) para precipitar cerca de la superficie de la Tierra, y la irradiación UV fue de 10 a 100 veces más intensa que ahora; de ahí que las propiedades fotosintéticas únicas mediadas por ZnS proporcionaran las condiciones energéticas adecuadas para energizar la síntesis de moléculas informativas y metabólicas y la selección de nucleobases fotoestables.
La teoría del mundo Zn se ha completado con evidencia experimental y teórica para la constitución iónica del interior de las primeras protocélulas antes de que evolucionaran las arqueas, las bacterias y los protoeucariotas. Archibald Macallum notó la semejanza de los fluidos corporales como la sangre y la linfa con el agua de mar; sin embargo, la composición inorgánica de todas las células difiere de la del agua marina moderna, lo que llevó a Mulkidjanian y sus colegas a reconstruir los "criaderos" de las primeras células combinando el análisis geoquímico con el escrutinio filogenómico de los requerimientos de iones inorgánicos de los componentes universales de las células modernas. Los autores concluyen que las proteínas y los sistemas funcionales omnipresentes, y por inferencia primordiales, muestran afinidad y requisitos funcionales para K, Zn, Mn y fosfato. La reconstrucción geoquímica muestra que la composición iónica propicia para el origen de las células no podría haber existido en lo que hoy llamamos entornos marinos, pero es compatible con las emisiones de zonas dominadas por vapor de lo que hoy llamamos sistemas geotérmicos tierra adentro. Bajo el oxígeno agotado, CO2 dominadas por la atmósfera primordial, la química de los condensados de agua y exhalaciones cerca de campos geotérmicos se asemejaría el medio interno de las células modernas. Por lo tanto, las etapas precelulares de la evolución pueden haber tenido lugar en "estanques Darwin" poco profundos revestidos con minerales de silicato poroso mezclados con sulfuros metálicos y enriquecidos en K, Zn y compuestos de fósforo.
Hipótesis de ventilación de mar profundo

Las rejillas de ventilación forman una fuente sostenida de energía química derivada de reacciones redox, en la cual los donadores de electrones (hidrógeno molecular) reaccionan con los aceptores de electrones (dióxido de carbono); ver teoría del mundo Hierro-azufre. Estas son reacciones altamente exotérmicas. Las rejillas de ventilación forman una fuente sostenida de energía química derivada de reacciones redox, en la cual los donadores de electrones (hidrógeno molecular) reaccionan con los aceptores de electrones (dióxido de carbono); ver teoría del mundo Hierro-azufre. Estas son reacciones altamente exotérmicas.
Michael Russell demostró que los conductos de ventilación alcalinos creaban un gradiente quiosmótico abiógeno de fuerza motriz protónica (PMF), en el que las condiciones son ideales para un criadero abiógeno de por vida. Sus compartimentos microscópicos "proporcionan un medio natural para concentrar moléculas orgánicas", compuestos de minerales de hierro y azufre como mackinawite, dotados de estas células minerales con las propiedades catalíticas previstas por Wächtershäuser. Este movimiento de iones a través de la membrana depende de una combinación de dos factores:
- La fuerza de difusión causada por el gradiente de concentración: todas las partículas, incluidos los iones, tienden a difundirse desde una concentración más alta a más baja.
- La fuerza electrostática causada por el gradiente de potencial eléctrico: los cationes como los protones H tienden a difundirse por el potencial eléctrico, los aniones en la dirección opuesta.
Estos dos gradientes tomados juntos se pueden expresar como un gradiente electroquímico, proporcionando energía para la síntesis abiogénica. La fuerza motriz del protón se puede describir como la medida de la energía potencial almacenada como una combinación de gradientes de protones y voltaje a través de una membrana (diferencias en la concentración de protones y el potencial eléctrico).
Jack W. Szostak sugirió que la actividad geotérmica proporciona mayores oportunidades para el origen de la vida en los lagos abiertos donde hay una acumulación de minerales. En 2010, basándose en el análisis espectral del mar y del agua mineral caliente, Ignat Ignatov y Oleg Mosin demostraron que la vida puede haberse originado predominantemente en agua mineral caliente. El agua mineral caliente que contiene bicarbonato e iones de calcio tiene el rango más óptimo. Este caso es similar al origen de la vida en respiraderos hidrotermales, pero con bicarbonato e iones de calcio en agua caliente. Esta agua tiene un pH de 9-11 y es posible tener las reacciones en agua de mar. De acuerdo con Melvin Calvin, ciertas reacciones de condensación-deshidratación de aminoácidos y nucleótidos en bloques individuales de péptidos y ácidos nucleicos pueden tener lugar en la hidrosfera primaria con pH 9-11 en una etapa evolutiva posterior. Algunos de estos compuestos como el ácido cianhídrico (HCN) han sido probados en los experimentos de Miller. Este es el entorno en el que se han creado los estromatolitos. David Ward de la Universidad Estatal de Montana describió la formación de estromatolitos en agua mineral caliente en el Parque Nacional Yellowstone. Los estromatolitos sobreviven en agua mineral caliente y cerca de áreas con actividad volcánica. Los procesos han evolucionado en el mar cerca de géiseres de agua mineral caliente. En 2011, Tadashi Sugawara de la Universidad de Tokio creó una protocélula en agua caliente. Algunos de estos compuestos como el ácido cianhídrico (HCN) han sido probados en los experimentos de Miller. Este es el entorno en el que se han creado los estromatolitos. David Ward de la Universidad Estatal de Montana describió la formación de estromatolitos en agua mineral caliente en el Parque Nacional Yellowstone. Los estromatolitos sobreviven en agua mineral caliente y cerca de áreas con actividad volcánica. Los procesos han evolucionado en el mar cerca de géiseres de agua mineral caliente. En 2011, Tadashi Sugawara de la Universidad de Tokio creó una protocélula en agua caliente. Algunos de estos compuestos como el ácido cianhídrico (HCN) han sido probados en los experimentos de Miller. Este es el entorno en el que se han creado los estromatolitos. David Ward de la Universidad Estatal de Montana describió la formación de estromatolitos en agua mineral caliente en el Parque Nacional Yellowstone. Los estromatolitos sobreviven en agua mineral caliente y cerca de áreas con actividad volcánica. Los procesos han evolucionado en el mar cerca de géiseres de agua mineral caliente. En 2011, Tadashi Sugawara de la Universidad de Tokio creó una protocélula en agua caliente. Los estromatolitos sobreviven en agua mineral caliente y cerca de áreas con actividad volcánica. Los procesos han evolucionado en el mar cerca de géiseres de agua mineral caliente. En 2011, Tadashi Sugawara de la Universidad de Tokio creó una protocélula en agua caliente. Los estromatolitos sobreviven en agua mineral caliente y cerca de áreas con actividad volcánica. Los procesos han evolucionado en el mar cerca de géiseres de agua mineral caliente. En 2011, Tadashi Sugawara de la Universidad de Tokio creó una protocélula en agua caliente.
La investigación experimental y el modelado computarizado sugieren que las superficies de las partículas minerales dentro de los respiraderos hidrotermales tienen propiedades catalíticas similares a las de las enzimas y son capaces de crear moléculas orgánicas simples, como metanol (CH 3 OH) y ácido fórmico, acético y pirúvico. disuelto CO 2 en el agua.
La investigación reportada arriba por William F. Martin en julio de 2016 apoya la tesis de que la vida surgió en los respiraderos hidrotermales, que la química espontánea en la corteza terrestre impulsada por interacciones roca-agua en el desequilibrio termodinámico sustentaron el origen de la vida y que los linajes fundadores de las arqueas y las bacterias eran autótrofos dependientes de H2 que usaban CO2 como su aceptor terminal en el metabolismo energético. Martin sugiere, en base a esta evidencia, que LUCA "pudo haber dependido en gran medida de la energía geotérmica del respiradero para sobrevivir".
Termosintesis
El proceso bioenergético actual de la fermentación se lleva a cabo mediante el ciclo del ácido cítrico antes mencionado o la vía del Acetil-CoA, que se han conectado al mundo primordial Hierro-azufre. En un enfoque diferente, la hipótesis de la termosíntesis considera el proceso bioenergético de la quimiosmosis, que desempeña un papel esencial en la respiración celular y la fotosíntesis, más basal que la fermentación: la enzima ATP sintasa, que sostiene la quimiosmosis, se propone como la enzima actualmente más estrechamente relacionada al primer proceso metabólico.
En primer lugar, la vida necesitaba una fuente de energía para producir la reacción de condensación que producía los enlaces peptídicos de las proteínas y los enlaces fosfodiéster del ARN. En una generalización y variación térmica del mecanismo de cambio de unión de la ATP sintasa de hoy en día, la "primera proteína" tendría sustratos unidos (péptidos, fosfato, nucleósidos, ARN 'monómeros') y los condensó a un producto de reacción que permaneció unido hasta después de una cambio de temperatura fue liberado por despliegue térmico.
La fuente de energía bajo la hipótesis de la termosíntesis era el ciclo térmico, el resultado de la suspensión de protocélulas en una corriente de convección, como es plausible en un manantial volcánico; la convección explica la autoorganización y la estructura disipativa requerida en cualquier origen del modelo de vida. El papel omnipresente del ciclo térmico en la germinación y la división celular se considera una reliquia de la termosíntesis primordial.
Al fosforilar los lípidos de la membrana celular, esta "primera proteína" dio una ventaja selectiva a la protocélula lipídica que contenía la proteína. Esta proteína también sintetizó una biblioteca de muchas proteínas, de las cuales solo una fracción diminuta tenía capacidades de termosíntesis. Según lo propuesto por Dyson, se propagó funcionalmente: creó hijas con capacidades similares, pero no se copió a sí misma. Las hijas funcionales consistían en diferentes secuencias de aminoácidos.
Mientras que el mundo Hierro-azufre identifica una vía circular como la más simple, la hipótesis de la termosíntesis ni siquiera invoca una vía: el mecanismo de cambio de unión de la ATP sintasa se asemeja a un proceso de adsorción física que produce energía libre, en lugar de un mecanismo enzimático regular, que disminuye la energía gratis. Se ha afirmado que la aparición de sistemas cíclicos de catalizadores de proteínas es inverosímil.
Otros modelos
Cronología de la naturaleza
-13 -
-
-12 -
-
-11 -
-
-10 -
-
-9 -
-
-8 -
-
-7 -
-
-6 -
-
-5 -
-
-4 -
-
-3 -
-
-2 -
-
-1 -
-
0 -
Escala del eje: mil millones de años
Hipótesis de arcilla
Montmorillonita , una arcilla abundante, es un catalizador para la polimerización del ARN y para la formación de membranas a partir de lípidos. Alexander Graham Cairns-Smith presentó en 1985 un modelo para el origen de la vida con arcilla y fue explorado como un mecanismo plausible por varios científicos. La hipótesis de la arcilla postula que las moléculas orgánicas complejas surgieron gradualmente en superficies de replicación no orgánicas preexistentes de cristales de silicato en solución.En el Instituto Politécnico Rensselaer, los estudios de James P. Ferris también han confirmado que los minerales arcillosos de montmorillonita catalizan la formación de ARN en solución acuosa, al unir nucleótidos para formar cadenas más largas.
En 2007, Bart Kahr de la Universidad de Washington y sus colegas informaron sobre sus experimentos que probaron la idea de que los cristales pueden actuar como fuente de información transferible, utilizando cristales de hidrogenoftalato de potasio. Los cristales de "madre" con imperfecciones se escindieron y se usaron como semillas para hacer crecer cristales "hijas" de la solución. Luego examinaron la distribución de imperfecciones en los nuevos cristales y encontraron que las imperfecciones en los cristales de la madre se reproducían en las hijas, pero los cristales hijas también tenían muchas imperfecciones adicionales. Para que se observe el comportamiento similar a un gen, la cantidad de herencia de estas imperfecciones debería haber excedido la de las mutaciones en las sucesivas generaciones, pero no fue así. Así Kahr concluyó que los cristales "
Modelo de "biosfera profunda" del oro
En la década de 1970, Thomas Gold propuso la teoría de que la vida no se desarrolló primero en la superficie de la Tierra, sino a varios kilómetros por debajo de la superficie. Se afirma que el descubrimiento de vida microbiana debajo de la superficie de otro cuerpo en nuestro sistema solar daría gran credibilidad a esta teoría. Thomas Gold también afirmó que se necesita un hilo de comida de una fuente profunda e inalcanzable para sobrevivir porque la vida que surge en un charco de materia orgánica es probable que consuma toda su comida y se extinga. La teoría del oro es que el flujo de tales alimentos se debe a la emisión de gases del metano primordial del manto de la Tierra;
Panspermia
Panspermia es la hipótesis de que la vida existe en todo el universo, distribuida por meteoritos, asteroides, cometas, planetoides y, también, por naves espaciales en forma de contaminación involuntaria por microorganismos.
La hipótesis de la panspermia no intenta explicar cómo se originó la vida, sino que simplemente la traslada a otro planeta o cometa. La ventaja de un origen extraterrestre de la vida primitiva es que no se requiere que la vida se haya formado en cada planeta en el que ocurre, sino en un solo lugar, y luego se disemina sobre la galaxia a otros sistemas estelares a través de un cometa y / o impacto de meteorito. La evidencia para respaldar la hipótesis es escasa, pero encuentra apoyo en estudios de meteoritos marcianos encontrados en la Antártida y en estudios de supervivencia de microbios extremófilos en pruebas en el espacio ultraterrestre. (Consulte también la lista de microorganismos probados en el espacio exterior).
Moléculas orgánicas extraterrestres
Un compuesto orgánico es cualquier miembro de una gran clase de sustancias químicas gaseosas, líquidas o sólidas cuyas moléculas contienen carbono. El carbono es el cuarto elemento más abundante en el Universo en masa después del hidrógeno, el helio y el oxígeno. El carbono es abundante en el Sol, las estrellas, los cometas y en las atmósferas de la mayoría de los planetas. Los compuestos orgánicos son relativamente comunes en el espacio, formados por "fábricas de síntesis molecular compleja" que se producen en nubes moleculares y envolventes circunestelares, y evolucionan químicamente después de que las reacciones se inician principalmente por radiación ionizante. Con base en estudios de modelos computarizados, las complejas moléculas orgánicas necesarias para la vida se pueden haber formado en los granos de polvo en el disco protoplanetario que rodea al Sol antes de la formación de la Tierra. De acuerdo con los estudios de computación,
Las observaciones sugieren que la mayoría de los compuestos orgánicos introducidos en la Tierra por partículas de polvo interestelar se consideran agentes principales en la formación de moléculas complejas, gracias a sus peculiares actividades catalíticas de superficie. Los estudios informados en 2008, basados en las relaciones isotópicas C / C de los compuestos orgánicos encontrados en el meteorito Murchison, sugirieron que el uracilo componente del ARN y las moléculas relacionadas, incluida la xantina, se formaron extraterrestre. El 8 de agosto de 2011, se publicó un informe basado en estudios de la NASA sobre los meteoritos encontrados en la Tierra que sugerían componentes de ADN (adenina, guanina y moléculas orgánicas relacionadas) en el espacio exterior. Los científicos también descubrieron que el polvo cósmico que permea el universo contiene compuestos orgánicos complejos ("sólidos orgánicos amorfos con una estructura mixto aromático-alifática"). ) que podría ser creado naturalmente, y rápidamente, por estrellas. Sun Kwok de la Universidad de Hong Kong sugirió que estos compuestos pueden haber estado relacionados con el desarrollo de la vida en la Tierra. Dijo que "si este es el caso, la vida en la Tierra podría haber tenido un tiempo más fácil para comenzar ya que estos orgánicos pueden servir como básicos ingredientes para la vida ".

Glycolaldehyde, el primer ejemplo de una molécula de azúcar interestelar, se detectó en la región de formación de estrellas cerca del centro de nuestra galaxia. Fue descubierto en 2000 por Jes Jørgensen y Jan M. Hollis. En 2012, el equipo de Jørgensen informó la detección de glicolaldehído en un sistema estelar distante. La molécula se encontró alrededor del binario protostelar IRAS 16293-2422 a 400 años luz de la Tierra. El glicocolaldehído es necesario para formar ARN, que es similar en función al ADN. Estos hallazgos sugieren que las moléculas orgánicas complejas se pueden formar en los sistemas estelares antes de la formación de los planetas, llegando eventualmente a los planetas jóvenes al inicio de su formación. Debido a que los azúcares están asociados con el metabolismo y el código genético, dos de los aspectos más básicos de la vida,
La NASA anunció en 2009 que los científicos habían identificado por primera vez otro elemento químico fundamental de la vida en un cometa, la glicina, un aminoácido que se detectó en material expulsado del cometa Wild 2 en 2004 y atrapado por Stardust de la NASA. sonda. La glicina se ha detectado en meteoritos antes. Carl Pilcher, quien dirige el Instituto de Astrobiología de la NASA, comentó que "el descubrimiento de la glicina en un cometa respalda la idea de que los componentes fundamentales de la vida prevalecen en el espacio y refuerza el argumento de que la vida en el universo puede ser común en lugar de rara. " Los cometas están incrustados con capas externas de material oscuro, que se cree que es una sustancia parecida al alquitrán compuesta de material orgánico complejo formado a partir de compuestos de carbono simples después de reacciones iniciadas principalmente por radiación ionizante. Es posible que una lluvia de material de los cometas pudiera haber traído cantidades significativas de tales moléculas orgánicas complejas a la Tierra. Los aminoácidos que se formaron extraterrestre también pueden haber llegado a la Tierra a través de cometas. Se estima que durante el Bombardeo Pesado Tardío,

Los hidrocarburos aromáticos policíclicos (HAP) son las moléculas poliatómicas más conocidas y abundantes en el universo observable, y se consideran un componente probable del mar primordial. En 2010, los HAP, junto con los fullerenos (o "buckyballs"), se han detectado en las nebulosas. En marzo de 2015, científicos de la NASA informaron que, por primera vez, se han formado en el laboratorio compuestos orgánicos complejos de vida como el uracilo, la citosina y la timina en condiciones de espacio ultraterrestre, utilizando sustancias químicas de partida, como la pirimidina, encontradas en meteoritos La pirimidina, como los HAP, la sustancia química más rica en carbono que se encuentra en el Universo, puede haberse formado en estrellas gigantes rojas o en polvo interestelar y nubes de gas.
Mundo de lípidos
La teoría del mundo lipídico postula que el primer objeto autorreplicante era lipídico. Se sabe que los fosfolípidos forman bicapas lipídicas en agua bajo agitación, la misma estructura que en las membranas celulares. Estas moléculas no estaban presentes en la Tierra primitiva, pero otras moléculas anfifílicas de cadena larga también forman membranas. Además, estos cuerpos pueden expandirse (mediante la inserción de lípidos adicionales), y bajo una expansión excesiva pueden sufrir una división espontánea que conserva el mismo tamaño y composición de lípidos en las dos progenies. La idea principal en esta teoría es que la composición molecular de los cuerpos lipídicos es la forma preliminar para el almacenamiento de la información, y la evolución condujo a la aparición de entidades de polímeros como ARN o ADN que pueden almacenar información favorablemente. Los estudios sobre vesículas de anfífilos potencialmente prebióticos se han limitado hasta ahora a los sistemas que contienen uno o dos tipos de anfífilos. Esto en contraste con la salida de reacciones químicas prebióticas simuladas, que típicamente producen mezclas muy heterogéneas de compuestos. Dentro de la hipótesis de una membrana de bicapa lipídica compuesta de una mezcla de diversos compuestos anfifílicos distintos existe la posibilidad de una gran cantidad de combinaciones teóricamente posibles en la disposición de estos anfífilos en la membrana. Entre todas estas posibles combinaciones, una disposición local específica de la membrana habría favorecido la constitución de un hiperciclo, en realidad una retroalimentación positiva compuesta por dos catalizadores mutuos representados por un sitio de membrana y un compuesto específico atrapado en la vesícula.
Polifosfatos
Un problema en la mayoría de los escenarios de abiogénesis es que el equilibrio termodinámico de los aminoácidos frente a los péptidos está en la dirección de los aminoácidos separados. Lo que se ha perdido es una fuerza que impulsa la polimerización. La resolución de este problema bien puede estar en las propiedades de los polifosfatos. Los polifosfatos se forman por polimerización de iones monofosfato ordinarios PO 4. Se han investigado varios mecanismos de síntesis de moléculas orgánicas. Los polifosfatos causan la polimerización de aminoácidos en péptidos. También son precursores lógicos en la síntesis de compuestos bioquímicos clave como el trifosfato de adenosina (ATP). Una cuestión clave parece ser que el calcio reacciona con el fosfato soluble para formar fosfato de calcio insoluble (apatita), por lo que debe encontrarse algún mecanismo plausible para evitar que los iones de calcio causen la precipitación de fosfato. Se ha trabajado mucho en este tema a lo largo de los años, pero una nueva idea interesante es que los meteoritos pueden haber introducido especies reactivas de fósforo en la Tierra primitiva.
Hipótesis del mundo PAH
Se sabe que los hidrocarburos aromáticos policíclicos (HAP) son abundantes en el universo, incluso en el medio interestelar, en los cometas y en los meteoritos, y son algunas de las moléculas más complejas que hasta ahora se han encontrado en el espacio.
Se han postulado otras fuentes de moléculas complejas, incluido el origen estelar o interestelar extraterrestre. Por ejemplo, a partir del análisis espectral, se sabe que las moléculas orgánicas están presentes en cometas y meteoritos. En 2004, un equipo detectó rastros de HAP en una nebulosa. En 2010, otro equipo también detectó HAP, junto con fullerenos, en nebulosas. El uso de HAP también se ha propuesto como un precursor del mundo del ARN en la hipótesis del mundo de HAP. El Telescopio Espacial Spitzer ha detectado una estrella, HH 46-IR, que se está formando por un proceso similar al que se formó el Sol. En el disco de material que rodea a la estrella, hay una gran variedad de moléculas, que incluyen compuestos de cianuro, hidrocarburos y monóxido de carbono. En septiembre de 2012, científicos de la NASA informaron que los HAP, sometidos a condiciones de medio interestelar, se transforman, a través de hidrogenación, oxigenación e hidroxilación, a compuestos orgánicos más complejos: "un paso en el camino hacia los aminoácidos y nucleótidos, las materias primas de las proteínas y el ADN, respectivamente". Además, como resultado de estas transformaciones, los HAP pierden su firma espectroscópica, lo que podría ser una de las razones "por la falta de detección de HAP en los granos de hielo interestelares, particularmente en las regiones externas de nubes frías y densas o en las capas moleculares superiores de los discos protoplanetarios "
La NASA mantiene una base de datos para rastrear los HAP en el universo. Más del 20% del carbono en el universo puede estar asociado con HAP, posibles materiales de partida para la formación de vida. Los PAH parecen haberse formado poco después del Big Bang, están diseminados por todo el universo y están asociados con nuevas estrellas y exoplanetas.
Hipótesis de playa radiactiva
Zachary Adam afirma que los procesos de las mareas que ocurrieron durante una época en la que la Luna estaba mucho más cerca pueden haber concentrado granos de uranio y otros elementos radiactivos en la marca de agua alta en las playas primordiales, donde pueden haber sido responsables de generar bloques de vida. Según los modelos de computadora, un depósito de tales materiales radiactivos podría mostrar la misma reacción nuclear autosostenida que la que se encuentra en la veta de mineral de uranio Oklo en Gabón. Dicha arena de playa radiactiva podría haber proporcionado suficiente energía para generar moléculas orgánicas, como aminoácidos y azúcares a partir de acetonitrilo en agua. El material de monazita radioactivo también ha liberado fosfato soluble en las regiones entre granos de arena, por lo que es biológicamente "accesible". Por lo tanto, aminoácidos, azúcares, y los fosfatos solubles podrían haber sido producidos simultáneamente, según Adam. Los actínidos radioactivos, abandonados en cierta concentración por la reacción, podrían haber formado parte de complejos organometálicos. Estos complejos podrían haber sido importantes catalizadores tempranos de los procesos vivos.
John Parnell ha sugerido que dicho proceso podría proporcionar parte del "crisol de la vida" en las primeras etapas de cualquier planeta rocoso húmedo temprano, siempre que el planeta sea lo suficientemente grande como para haber generado un sistema de placas tectónicas que aporta minerales radioactivos a la superficie. Como se cree que la Tierra primitiva tuvo muchas placas más pequeñas, podría haber proporcionado un entorno adecuado para tales procesos.
Disipación termodinámica
El físico austriaco Ludwig Boltzmann del siglo XIX reconoció por primera vez que la lucha por la existencia de organismos vivientes no era ni materia prima ni energía, sino que tenía que ver con la producción de entropía derivada de la conversión del espectro solar en calor por estos sistemas. Boltzmann se dio cuenta de que los sistemas vivos, como todos los procesos irreversibles, dependían de la disipación de un potencial químico generalizado para su existencia. En su libro “¿Qué es la vida”, el físico austríaco del siglo 20 Erwin Schrödinger hizo hincapié en la importancia de la visión profunda de Boltzmann en la naturaleza termodinámica irreversible de los sistemas vivos, lo que sugiere que esta fue la física y la química detrás del origen y evolución de la vida. Sin embargo, procesos irreversibles, y mucho menos sistemas vivos, no se pudo analizar convenientemente bajo esta perspectiva hasta que Lars Onsager, y más tarde Ilya Prigogine, desarrollaron un elegante formalismo matemático para tratar la "autoorganización" del material bajo un potencial químico generalizado. Este formalismo se conoció como Termodinámica clásica irreversible y Prigogine recibió el Premio Nobel de Química en 1977 "por sus contribuciones a la termodinámica no equilibrada, particularmente la teoría de las estructuras disipativas". El análisis de Prigogine demostró que si un sistema se dejaron evolucionar bajo un potencial externo impuesta, el material podría organizar espontáneamente (reducir su entropía) que forma lo que llamó “estructuras disipativas” lo que aumentaría la disipación del impuesto externamente potencial (aumentar el mundial producción de entropía).
En su "Teoría de la disipación termodinámica del origen y la evolución de la vida", Karo Michaelian ha tomado la visión de Boltzmann y el trabajo de Prigogine a sus últimas consecuencias con respecto al origen de la vida. Esta teoría postula que el sello distintivo del origen y la evolución de la vida es la estructura disipativa microscópica de los pigmentos orgánicos y su proliferación en toda la superficie de la Tierra. La vida actual aumenta la producción de entropía de la Tierra en su entorno solar mediante la disipación de los fotones ultravioleta y visible en el calor a través de pigmentos orgánicos en el agua. Este calor cataliza entonces una serie de procesos disipativos secundarias tales como las corrientes del ciclo del agua, el océano y viento, huracanes, etc. Michaelian argumenta que si la función termodinámica de la vida actual es producir entropía a través de la disipación de fotones en pigmentos orgánicos, entonces esta probablemente fue su función en sus comienzos. Resulta que tanto ARN como ADN cuando está en solución de agua son absorbentes muy fuertes y disipadores extremadamente rápidos de la luz ultravioleta dentro de la región de longitud de onda 230-290 nm (UV-C), que es una parte del espectro solar que podría haber penetrado en el atmósfera prebiótica De hecho, no solo el ARN y el ADN, sino muchas moléculas fundamentales de la vida (las comunes a los tres dominios de la vida) también son pigmentos que se absorben en el UV-C y muchos de ellos también tienen una afinidad química por el ARN y el ADN. Por lo tanto, los ácidos nucleicos pueden haber actuado como moléculas aceptoras de las moléculas donantes del pigmento de la antena excitada por fotones UV-C al proporcionar un canal ultrarrápido para la disipación. Michaelian ha demostrado utilizando el formalismo de la termodinámica irreversible no lineales que no habría existido durante el Archean un imperativo termodinámico para la síntesis fotoquímica UV-C abiogenic y la proliferación de estos pigmentos sobre toda la superficie de la Tierra si actuaban como catalizadores para aumentar la disipación de los fotones solares. Al final del Arcaico, con la luz UV-C que disipa la capa de ozono inducida por la vida en la atmósfera superior de la Tierra, habría sido cada vez más improbable que surgiera una vida completamente nueva que no dependiera del complejo rutas metabólicas ya existentes, ya que ahora la energía libre en los fotones que llegan a la superficie de la Tierra habría sido insuficiente para la ruptura directa y la reconstrucción de enlaces covalentes. Se ha sugerido, sin embargo,
Muchas características sobresalientes de las moléculas fundamentales de la vida (las que se encuentran en los tres dominios) apuntan directamente a la participación de la luz UV-C en la estructura disipativa de la vida incipiente. Algunos de los problemas más difíciles relacionados con el origen de la vida, como la replicación sin enzima de ARN y ADN, la homoquiralidad de las moléculas fundamentales y el origen de la codificación de información en ARN y ADN, también encuentran una explicación dentro del mismo marco termodinámico disipativo considerando la probable existencia de una relación entre la replicación primordial y la disipación de fotones UV-C. Michaelian sugiere que es erróneo esperar describir la emergencia, proliferación o incluso evolución de la vida sin una referencia abrumadora a la producción de entropía mediante la disipación de un potencial químico generalizado, en particular,
Génesis múltiple
Diferentes formas de vida con procesos de origen variable pueden haber aparecido casi simultáneamente en la historia temprana de la Tierra. Las otras formas pueden estar extintas (habiendo dejado fósiles distintivos a través de su diferente bioquímica, por ejemplo, tipos hipotéticos de bioquímica). Se ha propuesto que:
Piscinas hidrotermales fluctuantes en islas volcánicas o proto-continentes
Armid Mulkidjanian y sus coautores piensan que los ambientes marinos no proporcionaron el equilibrio iónico y la composición universalmente encontrados en las células, así como los iones requeridos por las proteínas esenciales y las ribozimas encontradas virtualmente en todos los organismos vivos, especialmente con respecto a la relación K / Na , Mn, Zn y concentraciones de fosfato. Los únicos entornos conocidos que imitan las condiciones necesarias en la Tierra se encuentran en piscinas hidrotermales terrestres alimentadas por conductos de vapor. Además, los depósitos minerales en estos ambientes bajo una atmósfera anóxica tienen un pH adecuado (en oposición a las piscinas actuales en una atmósfera oxigenada), contienen precipitados de sulfuro minerales que bloquean la radiación UV dañina, tienen ciclos de humectación / secado que concentran soluciones de sustrato a concentraciones susceptibles a la formación espontánea de polímeros de ácidos nucleicos, poliésteres y depsipéptidos, tanto por reacciones químicas en el entorno hidrotermal, como por la exposición a la luz UV durante el transporte desde los respiraderos a las piscinas adyacentes. Sus escenarios prebióticos hipotéticos son similares a los ambientes de ventilación oceánicos profundos más comúnmente hipotetizados, pero añaden componentes adicionales que ayudan a explicar las peculiaridades encontradas en las reconstrucciones del Último Ancestro Común Universal (LUCA) de todos los organismos vivos.
Bruce Damer y David Deamer han llegado a la conclusión de que las membranas celulares no se pueden formar en agua de mar salada y, por lo tanto, deben haberse originado en agua dulce. Antes de que se formaran los continentes, la única tierra seca en la Tierra serían las islas volcánicas, donde el agua de lluvia formaría lagunas donde los lípidos podrían formar las primeras etapas hacia las membranas celulares. Se supone que estos predecesores de células verdaderas se han comportado más como un superorganismo en lugar de estructuras individuales, donde las membranas porosas alojarían moléculas que se filtrarían y entrarían en otras protocélulas. Solo cuando las células verdaderas hayan evolucionado, se adaptarán gradualmente a ambientes más salinos y entrarán al océano.
Colín-García et al. (2016) discuten las ventajas y desventajas de los respiraderos hidrotérmicos como ambientes primitivos. Mencionan que las reacciones exergónicas en tales sistemas podrían haber sido una fuente de energía libre que promovió reacciones químicas, además de su alta diversidad mineralógica que implica la inducción de importantes gradientes químicos, favoreciendo así la interacción entre los donantes de electrones y los aceptores. Colín-García et al. (2016) también resumen un conjunto de experimentos propuestos para evaluar el papel de los respiraderos hidrotérmicos en la síntesis prebiótica.
Teoría de la información
Una teoría que habla sobre el origen de la vida en la Tierra y otros planetas rocosos plantea la vida como un sistema de información en el que el contenido de información crece debido a la selección. La vida debe comenzar con la mínima información posible, o una posible desviación mínima del equilibrio termodinámico, y requiere energía termodinámicamente libre accesible por medio de su contenido de información. Las circunstancias más benignas, las variaciones mínimas de entropía con abundante energía libre, sugieren el espacio poroso en los primeros kilómetros de la superficie. La energía libre se deriva de los productos condensados de las reacciones químicas que tienen lugar en la nebulosa de enfriamiento.
Obtenido de: https://en.wikipedia.org/wiki/Abiogenesis