Evolución humana
Definición

La evolución humana es el proceso evolutivo que condujo a la aparición de humanos anatómicamente modernos, comenzando con la historia evolutiva de los primates, en particular el género Homo , y llevando al surgimiento del Homo sapiens como una especie distinta de la familia homínida, los grandes simios. Este proceso implicó el desarrollo gradual de rasgos como el bipedismo humano y el lenguaje.
El estudio de la evolución humana involucra muchas disciplinas científicas, que incluyen antropología física, primatología, arqueología, paleontología, neurobiología, etología, lingüística, psicología evolutiva, embriología y genética. Los estudios genéticos muestran que los primates se separaron de otros mamíferos hace unos 85 millones de años , en el período Cretácico superior, y los primeros fósiles aparecen en el Paleoceno, hace aproximadamente 55 millones de años .
Dentro de la superfamilia Hominoidea (simios), la familia Hominidae se separó de la familia Hylobatidae (gibón) hace unos 15-20 millones de años; Los grandes simios africanos (subfamilia Homininae) divergieron de los orangutanes (Ponginae) hace unos 14 millones de años ; el Homininitribe (humanos, australopitecos y otros géneros bípedos extintos y chimpancés) se separó de la tribu Gorillini (gorilas) entre 8-9 millones de años atrás; y, a su vez, las subtribus Hominina (humanos y antepasados bípedos) y Panina (chimpancés) se separaron hace 4-7.5 millones de años.
Cambios anatómicos
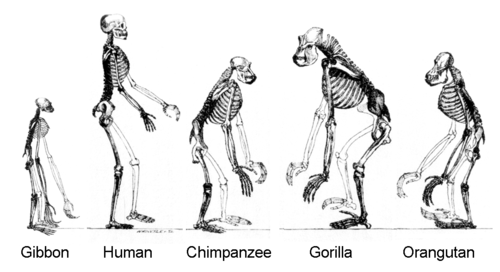
La evolución humana desde su primera separación del último ancestro común de humanos y chimpancés se caracteriza por una serie de cambios morfológicos, de desarrollo, fisiológicos y de comportamiento. Las más importantes de estas adaptaciones son bipedismo, aumento del tamaño del cerebro, ontogenia alargada (gestación e infancia) y disminución del dimorfismo sexual. La relación entre estos cambios es tema de debate continuo. Otros cambios morfológicos significativos incluyeron la evolución de un agarre de potencia y precisión, un cambio que se produjo por primera vez en H. erectus .
Bipedalismo
El bipedismo es la adaptación básica del homínido y se considera la causa principal detrás de un conjunto de cambios esqueléticos compartidos por todos los homínidos bípedos. El homínido más antiguo, de bipedismo supuestamente primitivo, se considera Sahelanthropus u Orrorin , ambos surgieron hace unos 6 a 7 millones de años. Los nudilleros no bípedos, el gorila y el chimpancé, divergieron de la línea de homínidos durante un período que abarca el mismo tiempo, por lo que cualquiera de Sahelanthropus u Orrorin puede ser nuestro último ancestro compartido. Ardipithecus , un bípedo completo, surgió un poco más tarde.
Los primeros bípedos eventualmente evolucionaron en los australopitecinos y aún más tarde en el género Homo. Existen varias teorías sobre el valor de adaptación del bipedismo. Es posible que se favoreciera el bipedismo porque liberaba las manos para alcanzar y transportar alimentos, ahorraba energía durante la locomoción, permitía correr y cazar a larga distancia, proporcionaba un campo de visión mejorado y ayudaba a evitar la hipertermia al reducir el área expuesta al sol directo ; tiene todas las ventajas para prosperar en el nuevo ambiente de sabana y bosques creado como resultado del levantamiento del Valle del Rift en el este de África en comparación con el hábitat forestal cerrado anterior. Un nuevo estudio respalda la hipótesis de que caminar sobre dos piernas, o bipedismo, evolucionó porque usaba menos energía que el movimiento de nudillo cuadrúpedo. Sin embargo, estudios recientes sugieren que la bipedestación sin la habilidad de usar fuego no habría permitido la dispersión global. Este cambio en la marcha produjo un alargamiento de las piernas proporcionalmente en comparación con la longitud de los brazos, que se acortaron mediante la eliminación de la necesidad de braquialismo. Otro cambio es la forma del dedo gordo del pie. Estudios recientes sugieren que los australopitecinos todavía vivieron parte del tiempo en los árboles como resultado de mantener el dedo gordo del pie agarrado. Esto se perdió progresivamente en Habilines.
Anatómicamente, la evolución del bipedismo se ha visto acompañada de una gran cantidad de cambios esqueléticos, no solo en las piernas y la pelvis, sino también en la columna vertebral, los pies y los tobillos, y el cráneo. El fémur evolucionó hacia una posición ligeramente más angular para mover el centro de gravedad hacia el centro geométrico del cuerpo. Las articulaciones de rodilla y tobillo se hicieron cada vez más robustas para soportar mejor el aumento de peso. Para soportar el aumento de peso en cada vértebra en posición vertical, la columna vertebral humana se convirtió en forma de S y las vértebras lumbares se hicieron más cortas y más anchas. En los pies, el dedo gordo se alineó con los otros dedos para ayudar a avanzar la locomoción. Los brazos y antebrazos se acortan en relación con las piernas, lo que facilita su funcionamiento. El foramen magnum migró debajo del cráneo y más anterior.
Los cambios más significativos se produjeron en la región pélvica, donde la hoja ilíaca que mira hacia abajo desde hace tiempo se acortó y se amplió como requisito para mantener estable el centro de gravedad mientras se camina; Los homínidos bípedos tienen una pelvis más corta, pero más ancha, similar a un cuenco debido a esto. Un inconveniente es que el canal de parto de los simios bípedos es más pequeño que en los simios que andan en nudillos, aunque ha habido un ensanchamiento de este en comparación con el australopitecino y los humanos modernos, permitiendo el paso de recién nacidos debido al aumento del tamaño craneal pero esto se limita a la parte superior, ya que un aumento adicional puede dificultar el movimiento bípedo normal.
El acortamiento de la pelvis y el canal de parto más pequeño evolucionó como un requisito para el bipedismo y tuvo efectos significativos en el proceso del nacimiento humano, que es mucho más difícil en los humanos modernos que en otros primates. Durante el nacimiento humano, debido a la variación en el tamaño de la región pélvica, la cabeza del feto debe estar en una posición transversal (en comparación con la madre) durante la entrada al canal de parto y girar unos 90 grados al salir. El canal de parto más pequeño se convirtió en un factor limitante del aumento del tamaño cerebral en los humanos primitivos y provocó un período de gestación más corto que conduce a la relativa inmadurez de la descendencia humana, que no puede caminar mucho antes de los 12 meses y tiene mayor neotenia, en comparación con otros primates. son móviles a una edad mucho más temprana. El aumento del crecimiento cerebral después del nacimiento y la mayor dependencia de los hijos de las madres tuvo un gran efecto en el ciclo reproductivo femenino, y la aparición más frecuente de alloparenting en los seres humanos en comparación con otros homínidos. La madurez sexual demorada también condujo a la evolución de la menopausia, con una explicación que establece que las mujeres mayores podrían transmitir mejor sus genes al cuidar de la descendencia de su hija, en comparación con tener más hijos propios.
Encefalización

La especie humana eventualmente desarrolló un cerebro mucho más grande que el de otros primates, típicamente 1,330 cm (81 cu) en humanos modernos, casi tres veces el tamaño de un chimpancé o cerebro de gorila. Después de un período de estasis con Australopithecus anamensis y Ardipithecus , especies que tenían cerebros más pequeños como resultado de su locomoción bípeda, el patrón de encefalización comenzó con el Homo habilis , cuyo cerebro de 600 cm era un poco más grande que el de los chimpancés. Esta evolución continuó en Homo erectus con 800-1.100 cm (49-67 pies cúbicos), y alcanzó un máximo en neandertales con 1.200-1.900 cm (73-116 pies cúbicos), incluso mayor que el Homo sapiens moderno. . Este aumento del cerebro se manifestó durante el crecimiento cerebral postnatal, excediendo con creces el de otros simios (heterocronía). También permitió largos períodos de aprendizaje social y adquisición del lenguaje en humanos juveniles, comenzando hace 2 millones de años.
Además, los cambios en la estructura de los cerebros humanos pueden ser aún más importantes que el aumento en el tamaño.

Los lóbulos temporales, que contienen centros para el procesamiento del lenguaje, han aumentado desproporcionadamente, al igual que la corteza prefrontal, que se ha relacionado con la toma de decisiones complejas y la moderación del comportamiento social. La encefalización se ha relacionado con un aumento de la carne y los almidones en la dieta. el desarrollo de la cocina, y se ha propuesto que la inteligencia aumentó como respuesta a una mayor necesidad de resolver problemas sociales a medida que la sociedad humana se hizo más compleja. Los cambios en la morfología del cráneo, como las mandíbulas más pequeñas y las conexiones de los músculos de la mandíbula, permitieron que el cerebro crezca más espacio.
El aumento en el volumen de la neocorteza también incluyó un rápido aumento en el tamaño del cerebelo. Su función se ha asociado tradicionalmente al equilibrio y al control motor fino, pero más recientemente con el habla y la cognición. Los grandes simios, incluidos los homínidos, tenían un cerebelo más pronunciado en relación con la neocorteza que otros primates. Se ha sugerido que debido a su función de control sensorio-motor y al aprendizaje de acciones musculares complejas, el cerebelo puede haber sustentado las adaptaciones tecnológicas humanas, incluidas las condiciones previas del habla.
La ventaja de supervivencia inmediata de la encefalización es difícil de discernir, ya que los principales cambios cerebrales del Homo erectus al Homo heidelbergensis no se acompañaron de cambios importantes en la tecnología. Se ha sugerido que los cambios fueron principalmente sociales y de comportamiento, incluyendo habilidades empáticas aumentadas, aumentos en el tamaño de los grupos sociales y una mayor plasticidad conductual
Dimorfismo sexual
El grado reducido de dimorfismo sexual en humanos es visible principalmente en la reducción del diente canino masculino en relación con otras especies de simios (excepto gibones) y crestas de cejas reducidas y la robustez general de los machos. Otro cambio fisiológico importante relacionado con la sexualidad en los humanos fue la evolución del estro oculto. Los seres humanos son los únicos homínidos en los que la hembra es fértil durante todo el año y en los que el cuerpo no produce señales especiales de fertilidad (como hinchazón genital o cambios manifiestos en la proceptividad durante el estro).
No obstante, los humanos conservan un grado de dimorfismo sexual en la distribución del vello corporal y la grasa subcutánea, y en el tamaño total, los machos son alrededor de un 15% más grandes que las hembras. Estos cambios tomados en conjunto han sido interpretados como resultado de un mayor énfasis en la vinculación de pares como una posible solución al requerimiento de una mayor inversión de los padres debido a la prolongada infancia de la descendencia.
Oposición cubana
La oposición cubital -el contacto entre el pulgar y la punta del dedo meñique de la misma mano- es exclusiva de los humanos anatómicamente modernos. En otros primates, el pulgar es corto e incapaz de tocar el dedo meñique. La oposición cubital facilita el agarre de precisión y el poder de agarre de la mano humana, lo que subyace en todas las manipulaciones hábiles.
Otros cambios
Varios otros cambios también han caracterizado la evolución de los humanos, entre ellos una mayor importancia en la visión más que en el olfato; un período de desarrollo juvenil más largo y una mayor dependencia infantil; un intestino más pequeño; metabolismo basal más rápido; pérdida de vello corporal; evolución de las glándulas sudoríparas; un cambio en la forma de la arcada dental de tener forma de U a ser parabólica; desarrollo de un mentón (encontrado en Homo sapiens solo); desarrollo de procesos estiloides; y el desarrollo de una laringe descendente.
Historia del estudio
Hominin Linea de tiempo
-10 -
-
-9 -
-
-8 -
-
-7 -
-
-6 -
-
-5 -
-
-4 -
-
-3 -
-
-2 -
-
-1 -
-
0 -
Escala del eje: millones de años
Antes de Darwin
La palabra homo , el nombre del género biológico al que pertenecen los humanos, es en latín "humano". Fue elegido originalmente por Carl Linnaeus en su sistema de clasificación. La palabra "humano" proviene del latín humanus , la forma adjetival del homo . El "homo" latino deriva de la raíz indoeuropea * dhghem , o "tierra". [50] Linneo y otros científicos de su época también consideraron a los grandes simios como los parientes más cercanos de los humanos en base a las similitudes morfológicas y anatómicas.
Darwin
La posibilidad de vincular a los seres humanos con los simios anteriores por descendencia se hizo evidente solo después de 1859 con la publicación de El origen de las especies , de Charles Darwin , en la que defendía la idea de la evolución de nuevas especies de las anteriores. El libro de Darwin no abordaba la cuestión de la evolución humana, y solo decía que "se arrojará luz sobre el origen del hombre y su historia".
Los primeros debates sobre la naturaleza de la evolución humana surgieron entre Thomas Henry Huxley y Richard Owen. Huxley abogó por la evolución humana de los simios al ilustrar muchas de las similitudes y diferencias entre los humanos y los simios, y lo hizo particularmente en su libro de 1863, Evidencia sobre el lugar del hombre en la naturaleza . Sin embargo, muchos de los primeros partidarios de Darwin (como Alfred Russel Wallace y Charles Lyell) inicialmente no estuvieron de acuerdo en que el origen de las capacidades mentales y las sensibilidades morales de los humanos pudiera explicarse por selección natural, aunque esto cambió más adelante. Darwin aplicó la teoría de la evolución y la selección sexual a los humanos cuando publicó The Descent of Man en 1871.
Primeros fósiles
Un problema importante en el siglo XIX fue la falta de intermediarios fósiles. Los restos de Neanderthal fueron descubiertos en una cantera de piedra caliza en 1856, tres años antes de la publicación de On the Origin of Species , y fósiles de Neanderthal habían sido descubiertos en Gibraltar incluso antes, pero originalmente se decía que eran restos humanos de una criatura que sufría algún tipo de la enfermedad A pesar del descubrimiento de Eugène Dubois en 1891 de lo que ahora se llama Homo erectus en Trinil, Java, fue solo en la década de 1920 cuando se descubrieron tales fósiles en África, que las especies intermedias comenzaron a acumularse. En 1925, Raymond Dart describió Australopithecus africanus. El espécimen tipo fue el Niño Taung, un niño australopitecino que fue descubierto en una cueva. Los restos del niño eran un pequeño cráneo notablemente bien conservado y un endocast del cerebro.
Aunque el cerebro era pequeño (410 cm), su forma era redondeada, a diferencia de los chimpancés y los gorilas, y más como un cerebro humano moderno. Además, el espécimen mostraba dientes caninos cortos, y la posición del foramen magnum (el agujero en el cráneo donde entra la espina dorsal) era evidencia de locomoción bípeda. Todos estos rasgos convencieron a Dart de que el Niño Taung era un ancestro humano bípedo, una forma de transición entre simios y humanos.
Los fósiles del este de África y Homo naledi en Sudáfrica

Durante las décadas de 1960 y 1970, se encontraron cientos de fósiles en el este de África en las regiones de Olduvai Gorge y Lake Turkana. La fuerza impulsora de estas búsquedas fue la familia Leakey, con Louis Leakey y su esposa Mary Leakey, y más tarde su hijo Richard y su nuera Meave, todos exitosos y reconocidos mundialmente cazadores de fósiles y paleoantropólogos. Desde los lechos fósiles de Olduvai y el lago Turkana, acumularon ejemplares de los primeros homínidos: los australopitecinos y las especies Homo , e incluso el Homo erectus .
Estos descubrimientos consolidaron a África como la cuna de la humanidad. A fines de la década de 1970 y en la década de 1980, Etiopía emergió como el nuevo punto caliente de la paleoantropología después de que "Lucy", el miembro fósil más completo de la especie Australopithecus afarensis , fuera encontrado en 1974 por Donald Johanson cerca de Hadar en la región desértica del Triángulo Afar del norte Etiopía. Aunque el espécimen tenía un cerebro pequeño, la pelvis y los huesos de las piernas eran casi idénticos en su función a los de los humanos modernos, lo que demuestra con certeza que estos homínidos se habían puesto en pie. Lucy fue clasificada como una nueva especie, Australopithecus afarensis , que se cree que está más relacionada con el género Homo como antepasado directo, o como pariente cercano de un ancestro desconocido, que cualquier otro homínido u hominino conocido de este rango de tiempo temprano; ver los términos "homínido" y "homínido". (El espécimen recibió el sobrenombre de "Lucy" después de la canción de los Beatles "Lucy in the Sky with Diamonds", que se tocó en voz alta y repetidamente en el campamento durante las excavaciones). El área del Afar Triangle revelaría más fósiles de homínidos, particularmente aquellos descubiertos o descritos por equipos encabezados por Tim D. White en la década de 1990, incluyendo Ardipithecus ramidus y Ardipithecus kadabba .
En 2013, se encontraron esqueletos fósiles de Homo naledi , una especie extinta de homínido asignada (provisionalmente) al género Homo , en el sistema Rising Star Cave, un sitio en la región sudafricana de Cradle of Humankind en la provincia de Gauteng, cerca de Johannesburgo. Hasta septiembre de 2015, fósiles de al menos quince individuos, que suman 1550 especímenes, han sido excavados en la cueva. La especie se caracteriza por una masa corporal y estatura similar a las poblaciones humanas de cuerpo pequeño, un volumen endocraneal más pequeño similar al Australopithecus y una morfología craneal (forma del cráneo) similar al Homo temprano especies. La anatomía esquelética combina características primitivas conocidas de los australopitecinos con características conocidas de los primeros homínidos. Las personas muestran signos de haber sido deliberadamente eliminados dentro de la cueva cerca del momento de la muerte. Los fósiles aún no han sido fechados.
La revolución genética

La revolución genética en los estudios de la evolución humana comenzó cuando Vincent Sarich y Allan Wilson midieron la fuerza de las reacciones inmunológicas cruzadas de la albúmina en suero sanguíneo entre pares de criaturas, incluidos los humanos y los simios africanos (chimpancés y gorilas). La fuerza de la reacción podría expresarse numéricamente como una distancia inmunológica, que a su vez era proporcional al número de diferencias de aminoácidos entre proteínas homólogas en diferentes especies. Al construir una curva de calibración de la ID de pares de especies con tiempos de divergencia conocidos en el registro fósil, los datos podrían usarse como un reloj molecular para estimar los tiempos de divergencia de pares con registros de fósiles más pobres o desconocidos.
En su artículo seminal de 1967 en Science , Sarich y Wilson calcularon el tiempo de divergencia de humanos y simios hace cuatro o cinco millones de años, en un momento en que las interpretaciones estándar del registro fósil daban esta divergencia de al menos 10 a 30 millones. años. Los descubrimientos posteriores de fósiles, especialmente "Lucy", y la reinterpretación de materiales fósiles más antiguos, en particular Ramapithecus , mostraron que las estimaciones más recientes son correctas y validaron el método de la albúmina.
El progreso en la secuenciación del ADN, específicamente el ADN mitocondrial (ADNmt) y luego el ADN del cromosoma Y (ADN-Y), hizo avanzar la comprensión de los orígenes humanos. La aplicación del principio del reloj molecular revolucionó el estudio de la evolución molecular.
Sobre la base de una separación del orangután entre hace 10 y 20 millones de años, estudios anteriores del reloj molecular sugirieron que había aproximadamente 76 mutaciones por generación que no fueron heredadas por los niños humanos de sus padres; esta evidencia apoyó el tiempo de divergencia entre los homínidos y los chimpancés señalados anteriormente. Sin embargo, un estudio de 2012 en Islandia de 78 niños y sus padres sugiere una tasa de mutación de solo 36 mutaciones por generación; este dato amplía la separación entre humanos y chimpancés a un período anterior de hace más de 7 millones de años (Ma). Investigaciones adicionales con 226 crías de poblaciones de chimpancés salvajes en 8 localidades sugieren que los chimpancés se reproducen a una edad promedio de 26.5 años; lo que sugiere que la divergencia humana de los chimpancés ocurrió hace entre 7 y 13 millones de años. Y estos datos sugieren que Ardipithecus (4.5 Ma), Orrorin (6 Ma) y Sahelanthropus (7 Ma) pueden estar en el linaje homínido, e incluso que la separación puede haber ocurrido fuera de la región del Rift de África Oriental.
Además, el análisis de los genes de las dos especies en 2006 proporciona evidencia de que después de que los ancestros humanos comenzaron a divergir de los chimpancés, el emparejamiento interespecie entre "protohumanos" y "proto-chimpancés" ocurrió con regularidad para cambiar ciertos genes en el nuevo gen piscina:
- Una nueva comparación de los genomas humano y de chimpancé sugiere que después de que los dos linajes se separaron, pueden haber comenzado el entrecruzamiento ... Un hallazgo principal es que los cromosomas X de los humanos y los chimpancés parecen haber divergido unos 1.2 millones de años más recientemente que los otros cromosomas.
La investigación sugiere:
- De hecho, hubo dos divisiones entre los linajes humano y chimpancé, seguido del primer cruce entre las dos poblaciones y luego una segunda división. La sugerencia de una hibridación ha sorprendido a los paleoantropólogos, que sin embargo están tratando los nuevos datos genéticos con seriedad.
La búsqueda de los primeros homínidos
En la década de 1990, varios equipos de paleoantropólogos trabajaban en toda África en busca de evidencia de la primera divergencia del linaje hominino de los grandes simios. En 1994, Meave Leakey descubrió Australopithecus anamensis . El hallazgo fue ensombrecido por el descubrimiento de Timd White en 1995 de Ardipithecus ramidus , que retrasó el registro fósil hace 4.2 millones de años .
En 2000, Martin Pickford y Brigitte Senut descubrieron, en las Colinas de Tugen de Kenia, un homínido bípedo de 6 millones de años al que denominaron Orrorin tugenensis . Y en 2001, un equipo dirigido por Michel Brunet descubrió el cráneo de Sahelanthropus tchadensis que databa de hace 7,2 millones de años , y que Brunet argumentó que era un bípedo, y por lo tanto un homínido, es decir, un homínido ( cf. Hominidae; términos "homínidos"). "y hominins).
Dispersión humana

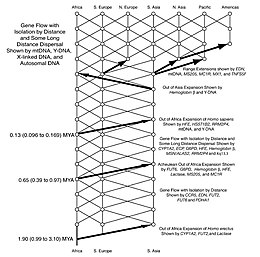
Los antropólogos en la década de 1980 se dividieron con respecto a algunos detalles de las barreras reproductivas y las dispersiones migratorias del género Homo . Posteriormente, la genética se ha utilizado para investigar y resolver estos problemas. Según la bomba del Sahara, la evidencia sugiere que el género Homo ha emigrado de África al menos tres o posiblemente cuatro veces (por ejemplo, Homo erectus , Homo heidelbergensis y dos o tres veces para el Homo sapiens ). La evidencia reciente sugiere que estas dispersiones están estrechamente relacionadas con los períodos fluctuantes del cambio climático.
La evidencia reciente sugiere que los humanos pueden haber abandonado África medio millón de años antes de lo que se pensaba. Un equipo conjunto franco-indio descubrió artefactos humanos en Siwalk Hills, al norte de Nueva Delhi, que se remontan al menos a 2,6 millones de años. Esto es anterior al hallazgo anterior más antiguo del género Homo en Dmanisi, en Georgia, que data de 1,85 millones de años. Aunque controvertido, las herramientas encontradas en una cueva china refuerzan el caso de que los humanos usaron herramientas hace 2.48 millones de años. Esto sugiere que la tradición asiática de la herramienta "Chopper", encontrada en Java y el norte de China puede haber dejado África antes de la aparición de el hacha de mano acheuliana.
Dispersión del Homo sapiens moderno
Hasta que la evidencia genética estuvo disponible hubo dos modelos dominantes para la dispersión de los humanos modernos. La hipótesis multirregional propuso que el género Homo contenía solo una sola población interconectada como lo hace hoy (no especies separadas), y que su evolución tuvo lugar en todo el mundo de forma continua en los últimos dos millones de años. Este modelo fue propuesto en 1988 por Milford H. Wolpoff. En contraste, el modelo "fuera de África" propuso que el H. sapiens moderno se especializó en África recientemente (es decir, hace aproximadamente 200,000 años) y la migración posterior a través de Eurasia resultó en el reemplazo casi completo de otras especies Homo . Este modelo ha sido desarrollado por Chris B. Stringer y Peter Andrews.
La secuenciación de mtDNA y Y-DNA muestreada de una amplia gama de poblaciones indígenas reveló información ancestral relacionada con el patrimonio genético masculino y femenino, y fortaleció la teoría de Out of Africa y debilitó los puntos de vista del Evolucionismo Multiregional. Alineados en las diferencias de árbol genético se interpretaron como un apoyo de un origen único reciente. Los análisis han demostrado una mayor diversidad de patrones de ADN en África, en consonancia con la idea de que África es el hogar ancestral de Eva mitocondrial y Adán cromosómico Y, y que la dispersión humana moderna fuera de África solo ha ocurrido en los últimos 55,000 años.
"Out of Africa" ha ganado mucho apoyo de la investigación que utiliza el ADN mitocondrial femenino y el cromosoma Y masculino. Después de analizar árboles genealógicos construidos con 133 tipos de ADN mt, los investigadores concluyeron que todos descienden de una progenitora africana femenina, apodada Eva mitocondrial. "Out of Africa" también se apoya en el hecho de que la diversidad genética mitocondrial es más alta entre las poblaciones africanas.
Un estudio amplio de la diversidad genética africana, dirigido por Sarah Tishkoff, encontró que la población San tenía la mayor diversidad genética entre las 113 poblaciones distintas de la muestra, lo que las convierte en uno de los 14 "grupos de población ancestrales". La investigación también localizó un posible origen de la migración humana moderna en el sudoeste de África, cerca de la frontera costera de Namibia y Angola. La evidencia fósil fue insuficiente para que el arqueólogo Richard Leakey resolviera el debate sobre dónde aparecieron por primera vez los humanos modernos en África. Los estudios de haplogrupos en ADN cromosómico Y y ADN mitocondrial han apoyado en gran medida un origen africano reciente. Toda la evidencia del ADN autosómico también es predominantemente compatible con un origen africano reciente. Sin embargo, hay evidencia de una mezcla arcaica en los humanos modernos, tanto en África como después,
La secuenciación reciente de los genomas de Neanderthal y Denisovan muestra que se ha producido cierta mezcla con estas poblaciones. Los humanos modernos fuera de África tienen 2-4% de alelos de Neanderthal en su genoma, y algunos melanesios tienen un 4-6% adicional de alelos de Denisovan. Estos nuevos resultados no contradicen el modelo "fuera de África", excepto en su interpretación más estricta, aunque hacen la situación más compleja. Después de la recuperación de un cuello de botella genético que posiblemente podría deberse a la catástrofe del supervolcán Toba, un grupo bastante pequeño salió de África y luego se cruzó brevemente en tres ocasiones con neandertales, probablemente en el medio oriente, en la estepa eurasiática o incluso en el norte de África antes de su partida. Sus descendientes aún predominantemente africanos se extendieron para poblar el mundo. Una fracción a su vez se cruzó con Denisovans, probablemente en el sudeste de Asia, antes de poblar Melanesia. Los haplotipos HLA de origen Neanderthal y Denisova se han identificado en poblaciones modernas de Eurasia y Oceanía. El gen Denisovan EPAS1 también se ha encontrado en poblaciones tibetanas.
Todavía hay diferentes teorías sobre si hubo un solo éxodo de África o varios. Un modelo de dispersión múltiple implica la teoría de Dispersión del Sur, que ha ganado apoyo en los últimos años a partir de evidencia genética, lingüística y arqueológica. En esta teoría, hubo una dispersión costera de humanos modernos desde el Cuerno de África cruzando el Bab el Mandib hacia Yemen a un nivel del mar más bajo hace unos 70,000 años. Este grupo ayudó a poblar el sudeste de Asia y Oceanía, explicando el descubrimiento de los primeros sitios humanos en estas áreas mucho antes que en el Levante. Este grupo parece haber dependido de los recursos marinos para su supervivencia.
Stephen Oppenheimer ha propuesto una segunda ola de humanos que luego se haya dispersado a través de los oasis del Golfo Pérsico y las montañas Zagros hacia el Medio Oriente. Alternativamente, puede haber atravesado la Península del Sinaí en Asia, desde poco después de 50,000 años BP, resultando en la mayor parte de las poblaciones humanas de Eurasia. Se ha sugerido que este segundo grupo posiblemente poseía una tecnología más sofisticada de "caza mayor" y dependía menos de las fuentes de alimentos costeros que el grupo original. Gran parte de la evidencia de la expansión del primer grupo habría sido destruida por el aumento del nivel del mar al final de cada máximo glacial.
Stephen Oppenheimer, sobre la base de la fecha temprana de Aurignacian iraní de Badoshan, sugiere que esta segunda dispersión pudo haber ocurrido con un período pluvial de unos 50,000 años antes del presente, con modernas culturas de cacería humanas extendiéndose por las montañas Zagros, llevando modernos genomas humanos de Omán, en todo el Golfo Pérsico, hacia el norte en Armenia y Anatolia, con una variante que viaja al sur hacia Israel y hacia Cyrenicia.
Evidencia
La evidencia sobre la cual se basan los relatos científicos de la evolución humana proviene de muchos campos de la ciencia natural. La principal fuente de conocimiento sobre el proceso evolutivo ha sido tradicionalmente el registro fósil, pero desde el desarrollo de la genética a partir de la década de 1970, el análisis del ADN ha llegado a ocupar un lugar de importancia comparable. Los estudios de ontogenia, filogenia y especialmente la biología evolutiva evolutiva de vertebrados e invertebrados ofrecen una visión considerable de la evolución de toda la vida, incluida la evolución de los humanos. El estudio específico del origen y la vida de los humanos es la antropología, particularmente la paleoantropología, que se centra en el estudio de la prehistoria humana.
Evidencia de la biología molecular

Los parientes vivos más cercanos de los humanos son los bonobos y los chimpancés (ambos del género Pan ) y los gorilas (el género Gorilla ). Con la secuenciación del genoma humano y del chimpancé, a partir de 2012 las estimaciones de la similitud entre sus secuencias de ADN oscilan entre el 95% y el 99%. Al usar la técnica llamada reloj molecular que estima el tiempo requerido para que se acumule la cantidad de mutaciones divergentes entre dos linajes, se puede calcular la fecha aproximada para la división entre linajes.
Los gibones (familia Hylobatidae) y luego los orangutanes (género Pongo ) fueron los primeros grupos en separarse de la línea que conduce a los homínidos, incluidos los humanos, seguidos por los gorilas y, finalmente, por los chimpancés (género Pan ). La fecha de división entre los linajes de los homínidos y los chimpancés es colocada por algunos hace entre 4 y 8 millones de años , es decir, durante el Mioceno superior. La especiación, sin embargo, parece haber sido inusualmente prolongada. La divergencia inicial se produjo en algún momento entre hace 7 a 13 millones de años , pero la hibridación en curso empañó la separación y retrasó la separación completa durante varios millones de años. Patterson (2006) fecha la divergencia final hace 5 a 6 millones de años .
La evidencia genética también se ha empleado para resolver la cuestión de si había algún flujo de genes entre los primeros humanos modernos y los neandertales, y para mejorar nuestra comprensión de los primeros patrones de migración humana y las fechas de división. Al comparar las partes del genoma que no están bajo selección natural y que, por lo tanto, acumulan mutaciones a un ritmo bastante constante, es posible reconstruir un árbol genético que incorpore toda la especie humana desde el último ancestro compartido.
Cada vez que aparece una cierta mutación (polimorfismo de un solo nucleótido) en un individuo y se transmite a sus descendientes, se forma un haplogrupo que incluye a todos los descendientes del individuo que también portarán esa mutación. Al comparar el ADN mitocondrial que se hereda solo de la madre, los genetistas han llegado a la conclusión de que el último ancestro común femenino cuyo marcador genético se encuentra en todos los humanos modernos, la llamada Eva mitocondrial, debe haber vivido hace unos 200,000 años.
Genética
La genética evolutiva humana estudia cómo un genoma humano difiere del otro, el pasado evolutivo que lo originó y sus efectos actuales. Las diferencias entre los genomas tienen implicaciones y aplicaciones antropológicas, médicas y forenses. Los datos genéticos pueden proporcionar información importante sobre la evolución humana.
Evidencia del registro fósil


Hay poca evidencia fósil de la divergencia de los linajes de los gorilas, los chimpancés y los homínidos. Los primeros fósiles que se han propuesto como miembros del linaje de los homínidos son Sahelanthropus tchadensis, que datan de hace 7 millones de años , Orrorin tugenensis, que data de hace 5,7 millones de años , y Ardipithecus kadabba, que data de hace 5,6 millones de años . Se ha argumentado que cada uno de estos es un antepasado bípedo de los homínidos posteriores, pero, en cada caso, las afirmaciones han sido impugnadas. También es posible que una o más de estas especies sean ancestros de otra rama de los simios africanos, o que representen un ancestro compartido entre los homínidos y otros simios.
La cuestión entonces de la relación entre estas especies fósiles tempranas y el linaje de los homínidos aún está por resolverse. De estas especies tempranas, los australopitecinos surgieron hace unos 4 millones de años y divergieron en ramas robustas (también llamadas Paranthropus ) y gráciles, una de las cuales (posiblemente A. garhi ) probablemente pasó a ser antepasados del género Homo . La especie australopitecina mejor representada en el registro fósil es Australopithecus afarensis con más de cien fósiles representados, encontrados desde el norte de Etiopía (como la famosa "Lucy"), hasta Kenia y Sudáfrica. Fósiles de australopitecos robustos como Au. robustus (o alternativamente Paranthropus robustus ) y Au./P. boisei son particularmente abundantes en Sudáfrica en sitios como Kromdraai y Swartkrans, y alrededor del lago Turkana en Kenia.
El miembro más antiguo del género Homo es el Homo habilis, que se desarrolló hace unos 2,8 millones de años . Homo habilis es la primera especie para la cual tenemos evidencia positiva del uso de herramientas de piedra. Desarrollaron la tecnología lítica Oldowan, llamada así por la Garganta de Olduvai en la que se encontraron los primeros especímenes. Algunos científicos consideran que el Homo rudolfensis , un grupo de fósiles más grandes con una morfología similar a los fósiles originales de H. habilis , es una especie separada, mientras que otros consideran que forman parte de H. habilis.-Simplemente representando variación intraespecífica, o tal vez incluso dimorfismo sexual. Los cerebros de estos primeros homínidos eran aproximadamente del mismo tamaño que los de un chimpancé, y su principal adaptación fue el bipedismo como una adaptación a la vida terrestre.
Durante el próximo millón de años, comenzó un proceso de encefalización y, por la llegada (hace aproximadamente 1.9 millones de años ) de Homo erectus en el registro fósil, la capacidad craneana se había duplicado. Homo erectus fue el primero de los homínidos en emigrar de África, y, hace 1,8 a 1,3 millones de años , esta especie se extendió a través de África, Asia y Europa. Una población de H. erectus , también a veces clasificada como una especie separada de Homo ergaster , permaneció en África y evolucionó hacia Homo sapiens . Se cree que estas especies, H. erectus y H. ergaster , fueron las primeras en usar fuego y herramientas complejas.
Los primeros fósiles de transición entre H. ergaster / erectus y H. sapiens arcaico son de África, como Homo rhodesiensis , pero también se encontraron formas aparentemente transitorias en Dmanisi, Georgia. Estos descendientes de H. erectus africano se diseminaron a través de Eurasia desde ca. Hace 500,000 años evolucionando en H. antecessor , H. heidelbergensis y H. neanderthalensis . Los primeros fósiles de humanos anatómicamente modernos son del Paleolítico Medio, hace unos 200,000 años, como los restos Omo de Etiopía; fósiles posteriores de la cueva de Es Skhul en Israel y el sur de Europa comenzaron hace unos 90,000 años (hace 0.09 millones de años ).
A medida que los humanos modernos se dispersaron desde África, se encontraron con otros homínidos como Homo neanderthalensis y los llamados Denisovans, que pueden haber evolucionado a partir de poblaciones de Homo erectus que habían abandonado África hace unos 2 millones de años . La naturaleza de la interacción entre los primeros humanos y estas especies hermanas ha sido una fuente de controversia de larga data, la cuestión es si los humanos reemplazaron estas especies anteriores o si de hecho eran lo suficientemente similares como para cruzarse, en cuyo caso estas poblaciones anteriores pueden haber contribuido material genético a los humanos modernos.
Se estima que esta migración fuera de África comenzó hace unos 70,000 años BP y los humanos modernos posteriormente se expandieron globalmente, reemplazando a los homínidos anteriores ya sea a través de la competencia o la hibridación. Habitaron Eurasia y Oceanía por 40,000 años BP, y las Américas por al menos 14,500 años BP.
Antes de Homo
La evolución temprana de los primates
La historia evolutiva de los primates se remonta a 65 millones de años. Una de las especies de mamíferos más antiguas conocidas como primates, Plesiadapis , vino de América del Norte; otro, Archicebus , vino de China. Otros primates basales similares se extendieron en Eurasia y África durante las condiciones tropicales del Paleoceno y Eoceno.

David R. Begun concluyó que los primeros primates florecieron en Eurasia y que un linaje que conduce a los simios y humanos africanos, incluido Dryopithecus , emigró al sur de Europa o Asia occidental hacia África. La población tropical superviviente de primates -que se ve más completamente en el Eoceno superior y en los lechos fósiles del Oligoceno inferior de la depresión Faiyum al suroeste de El Cairo- dio origen a todas las especies de primates existentes, incluidos los lémures de Madagascar, los loris del sudeste asiático, galagos o " Bush bebés "de África, y para los antropoides, que son los Platyrrhines o los monos del Nuevo Mundo, los Catarrhines o monos del Viejo Mundo, y los grandes simios, incluidos los humanos y otros homínidos.
El primer catarrino conocido es Kamoyapithecus del Oligoceno superior en Eragaleit en el norteño Gran Valle del Rift en Kenia, fechado hace 24 millones de años. Se cree que su ascendencia es una especie relacionada con Aegyptopithecus , Propliopithecus y Parapithecus del Faiyum, hace alrededor de 35 millones de años. En 2010, Saadanius fue descrito como un pariente cercano del último ancestro común de los catarrinos de la corona, y provisionalmente data de hace 29-28 millones de años, ayudando a llenar una brecha de 11 millones de años en el registro fósil.

En el Mioceno temprano, hace aproximadamente 22 millones de años, los muchos tipos de catarrinos primitivos adaptados arboreally del este de África sugieren una larga historia de diversificación previa. Los fósiles de hace 20 millones de años incluyen fragmentos atribuidos a Victoriapithecus , el primer mono del Viejo Mundo. Entre los géneros que se consideran en el linaje de los simios que conduce hasta hace 13 millones de años son Procónsul , rangwapithecus , dendropithecus , Limnopithecus , nacholapithecus , Equatorius , Nyanzapithecus, Afropithecus , heliopithecus y Kenyapithecus , todo ello desde el este de África.
La presencia de otros no cercopithecids generalizadas del Mioceno Medio de sitios ahora distant- Otavipithecus de los depósitos de la cueva en Namibia y Pierolapithecus y Dryopithecus de Francia, España y Austria-evidencia de una amplia diversidad de formas a través de África y la cuenca mediterránea durante el regímenes climáticos relativamente cálidos y equitativos del Mioceno temprano y medio. El más joven de los hominoides del Mioceno, Oreopithecus , proviene de lechos de carbón en Italia que datan de hace 9 millones de años.
La evidencia molecular indica que el linaje de gibones (familia Hylobatidae) divergió de la línea de los grandes simios hace unos 18-12 millones de años, y el de los orangutanes (subfamilia Ponginae) se separó de los otros grandes simios a los 12 millones de años; no hay fósiles que documenten claramente la ascendencia de los gibones, que pueden haberse originado en una población de homínidos del sudeste asiático hasta el momento desconocida, pero los proto-orangutanes fósiles pueden estar representados por Sivapithecus de la India y Griphopithecus de Turquía, de alrededor de 10 Hace millones de años.
Divergencia del clado humano de otros grandes simios

Las especies cercanas al último ancestro común de gorilas, chimpancés y humanos pueden estar representadas por fósiles de Nakalipithecus encontrados en Kenia y Ouranopithecus encontrados en Grecia. La evidencia molecular sugiere que hace entre 8 y 4 millones de años, primero los gorilas y luego los chimpancés (género Pan ) se separaron de la línea que conduce a los humanos. El ADN humano es aproximadamente 98.4% idéntico al de los chimpancés cuando se comparan polimorfismos de un solo nucleótido (ver genética evolutiva humana). El registro fósil, sin embargo, de gorilas y chimpancés es limitado; Ambos suelos de preservación de bosques lluviosos tienden a ser ácidos y disolver los huesos, y el sesgo de muestreo probablemente contribuya a este problema.
Otros homínidos probablemente se adaptaron a los ambientes más secos fuera del cinturón ecuatorial; y allí encontraron antílopes, hienas, perros, cerdos, elefantes, caballos y otros. El cinturón ecuatorial se contrajo después de unos 8 millones de años atrás, y hay muy poca evidencia fósil de la división-pensamiento que ocurrió en ese momento-del linaje de los homínidos de los linajes de los gorilas y los chimpancés. Los primeros fósiles argumentados por algunos que pertenecen al linaje humano son Sahelanthropus tchadensis (7 Ma) y Orrorin tugenensis (6 Ma), seguidos por Ardipithecus (5.5-4.4 Ma), con la especie Ar. kadabba y Ar. Ramidus .
Se ha argumentado en un estudio de la historia de la vida de Ar. Ramidus que la especie proporciona evidencia de un conjunto de adaptaciones anatómicas y de comportamiento en los primeros homínidos a diferencia de cualquier especie de gran simio existente. Este estudio demostró afinidades entre la morfología del cráneo de Ar. ramidus y el de los chimpancés lactantes y juveniles, lo que sugiere que la especie evolucionó a una morfología craneofacial juvenil o pedomórfica a través de la disociación heterocrónica de las trayectorias de crecimiento. También se argumentó que la especie proporciona apoyo para la noción de que los homininos muy temprano, similar a los bonobos ( Pan paniscus)) las especies de chimpancés menos agresivas, pueden haber evolucionado a través del proceso de autodomesticación. En consecuencia, argumentando en contra del llamado "modelo referencial de los chimpancés", los autores sugieren que ya no es sostenible el uso de comportamientos sociales y de apareamiento de los chimpancés comunes ( Pan troglodytes ) en los modelos de la evolución social de los primeros homininos. Al comentar sobre la ausencia de morfología canina agresiva en Ar. Ramidus y las implicaciones que esto tiene para la evolución de la psicología social de los homínidos, escribieron:
Los autores argumentan que muchas de las adaptaciones humanas básicas evolucionaron en los bosques antiguos y los ecosistemas de bosques de finales del Mioceno y principios del Plioceno de África. En consecuencia, argumentan que los humanos pueden no representar la evolución de un ancestro similar a un chimpancé como se ha supuesto tradicionalmente. Esto sugiere que muchas adaptaciones humanas modernas representan rasgos filogenéticamente profundos y que el comportamiento y la morfología de los chimpancés pueden haber evolucionado después de la división con el ancestro común que comparten con los humanos.
Género Australopithecus
El género Australopithecus evolucionó en el este de África hace unos 4 millones de años antes de extenderse por todo el continente y extinguirse hace 2 millones de años. Durante este período de tiempo existieron varias formas de australopiths, incluyendo Australopithecus anamensis , Au. afarensis , Au. sediba , y Au. africanus . Todavía existe cierto debate entre los académicos sobre si ciertas especies de homínidos africanos de esta época, como Au. robustus y Au. boisei , constituyen miembros del mismo género; si es así, se considerarían Au. Australopiths robustos mientras que los otros se considerarían Au. australopitos graciles. Sin embargo, si estas especies realmente constituyen su propio género, entonces se les puede dar su propio nombre, el Paranthropus .
- Australopithecus (4-1.8 Ma), con especies Au. Anamensis , Au. afarensis , Au. africanus , Au. bahrelghazali , Au. garhi y Au. sediba ;
- Kenyanthropus (3-2.7 Ma), con la especie K. platyops ;
- Paranthropus (3-1.2 Ma), con las especies P. aethiopicus , P. boisei y P. robustus
Se afirma que una nueva especie propuesta, Australopithecus deyiremeda, fue descubierta viviendo en el mismo período de Au. afarensis . Hay debate si Au.deyiremeda es una especie nueva o es Au. afarensis. El Australopithecus prometheus , también conocido como Little Foot, se ha fechado recientemente con 3,67 millones de años mediante una nueva técnica de datación, lo que hace que el género Australopithecus sea tan antiguo como el afarensis . Dado el dedo gordo oponible encontrado en Little Foot, parece que fue un buen escalador, y se cree que dado los depredadores nocturnos de la región, probablemente, como los gorilas y los chimpancés, construyó una plataforma de anidación en la noche, en los árboles.
Evolución del género Homo
El primer representante documentado del género Homo es el Homo habilis , que se desarrolló hace unos 2.8 millones de años , y es posiblemente la especie más temprana para la cual hay evidencia positiva del uso de herramientas de piedra. Los cerebros de estos primeros homínidos eran aproximadamente del mismo tamaño que los de un chimpancé, aunque se ha sugerido que este fue el momento en el que el gen humano SRGAP2 se duplicó, produciendo un cableado más rápido de la corteza frontal. Durante el próximo millón de años se produjo un proceso de encefalización rápida, y con la llegada de Homo erectus y Homo ergaster en el registro fósil, la capacidad craneana se había duplicado a 850 cm. (Tal aumento en el tamaño del cerebro humano es equivalente a que cada generación tenga 125,000 neuronas más que sus padres.) Se cree que Homo erectus y Homo ergaster fueron los primeros en usar el fuego y herramientas complejas, y fueron las primeras de la línea de homínidos en abandonar África, extendiéndose por África, Asia y Europa hace entre 1,3 y 1,8 millones de años .
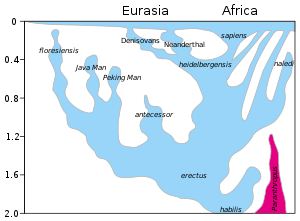

Según el reciente origen africano de la teoría de los humanos modernos, los humanos modernos evolucionaron en África posiblemente a partir de Homo heidelbergensis , Homo rhodesiensis u Homo antecessor y emigraron del continente hace unos 50,000 a 100,000 años, reemplazando gradualmente a las poblaciones locales de Homo erectus , Denisova hominins, Homo floresiensis y Homo neanderthalensis . Homo sapiens arcaico , el precursor de humanos anatómicamente modernos, evolucionó en el Paleolítico Medio entre 400,000 y 250,000 años atrás. La evidencia reciente de DNA sugiere que varios haplotipos de origen neandertal están presentes en todas las poblaciones no africanas, y los neandertales y otros homininos, como los denisovanos, pueden haber contribuido hasta el 6% de su genoma a los humanos actuales, lo que sugiere una interferencia limitada. crianza entre estas especies. La transición a la modernidad del comportamiento con el desarrollo de la cultura simbólica, el lenguaje y la tecnología lítica especializada ocurrió hace unos 50,000 años según algunos antropólogos, aunque otros apuntan a la evidencia que sugiere que un cambio gradual en el comportamiento tuvo lugar durante un lapso de tiempo más largo.
Homo sapiens es la única especie existente de su género, Homo . Mientras que algunas (extintas) especies de Homo podrían haber sido antepasados del Homo sapiens , muchos, tal vez la mayoría, eran probablemente "primos", habiéndose alejado de la línea ancestral de los homínidos. Todavía no hay consenso sobre cuáles de estos grupos deberían considerarse como una especie separada y cuáles deberían ser una subespecie; esto puede deberse a la escasez de fósiles oa las leves diferencias utilizadas para clasificar especies en el género Homo . La teoría de la bomba del Sahara (que describe un desierto del Sáhara "húmedo" en ocasiones aceptable) ofrece una posible explicación de la variación temprana en el género Homo .
Con base en la evidencia arqueológica y paleontológica, ha sido posible inferir, en cierta medida, las antiguas prácticas dietéticas de varias especies de Homo y estudiar el papel de la dieta en la evolución física y conductual dentro de Homo .
Algunos antropólogos y arqueólogos suscriben la teoría de la catástrofe Toba, que postula que la superupción del lago Toba en la isla de Sumatra en Indonesia hace unos 70,000 años causó consecuencias globales, matando a la mayoría de los humanos y creando un cuello de botella que afectó la herencia genética de todos los humanos hoy.
H. habilis y H. gautengensis
El Homo habilis vivió entre 2.8 y 1.4 Ma. La especie evolucionó en el sur y el este de África en el Plioceno superior o en el Pleistoceno temprano, 2.5-2 Ma, cuando divergió de los australopitecinos. Homo habilis tenía molares más pequeños y cerebros más grandes que los australopitecinos, e hizo herramientas de piedra y quizás huesos de animales. Uno de los primeros homínidos conocidos fue apodado 'handy man' por el descubridor Louis Leakey debido a su asociación con herramientas de piedra. Algunos científicos han propuesto trasladar esta especie de Homo a Australopithecus debido a que la morfología de su esqueleto está más adaptada a vivir en los árboles que a moverse sobre dos patas, como el Homo sapiens .
En mayo de 2010, se descubrió una nueva especie, Homo gautengensis , en Sudáfrica.
H. rudolfensis y H. georgicus
Estos son nombres de especies propuestos para fósiles de aproximadamente 1.9-1.6 Ma, cuya relación con el Homo habilis aún no está clara.
- Homo rudolfensis se refiere a un solo cráneo incompleto de Kenia. Los científicos han sugerido que este era otro Homo habilis , pero esto no ha sido confirmado.
- Homo georgicus , de Georgia, puede ser una forma intermedia entre Homo habilis y Homo erectus , o una subespecie de Homo erectus .
H. ergaster y H. erectus
Los primeros fósiles de Homo erectus fueron descubiertos por el médico holandés Eugene Dubois en 1891 en la isla indonesia de Java. Originalmente nombró al material Anthropopithecus erectus (1892-1893, considerado un punto como un primate fósil similar a un chimpancé) y Pithecanthropus erectus (1893-1894), cambiando su forma de pensar en función de su morfología, que consideró intermedia entre ese de humanos y simios). Años más tarde, en el siglo XX, el médico y paleoantropólogo alemán Franz Weidenreich (1873-1948) comparó en detalle los personajes del Hombre Java de Dubois, entonces llamado Pithecanthropus erectus , con los personajes del Hombre de Pekín, entonces llamado Sinanthropus pekinensis.. Weidenreich concluyó en 1940 que, debido a su similitud anatómica con los humanos modernos, era necesario reunir todos estos especímenes de Java y China en una sola especie del género Homo , la especie Homo erectus. Homo erectus vivió desde alrededor de 1.8 Ma hasta hace unos 70,000 años, lo que indicaría que probablemente fueron aniquilados por la catástrofe de Toba; sin embargo, Homo floresiensiscercano sobrevivió. La primera fase de Homo erectus , de 1.8 a 1.25 Ma, es considerada por algunos como una especie separada, Homo ergaster , o como Homo erectus ergaster , una subespecie de Homo erectus .
En África en el Pleistoceno temprano, 1.5-1 Ma, se cree que algunas poblaciones de Homo habilishan desarrollado cerebros más grandes y han fabricado herramientas de piedra más elaboradas; estas diferencias y otras son suficientes para que los antropólogos las clasifiquen como una nueva especie, Homo erectus-en África. Se cree que la evolución de las rodillas bloqueadas y el movimiento del foramen magnum son los posibles impulsores de los grandes cambios en la población. Esta especie también puede haber usado fuego para cocinar carne. Richard Wrangham sugiere que el hecho de que Homo parece haber sido una vivienda molida, con una longitud intestinal reducida, una dentición más pequeña, "y aumentó nuestros cerebros a su tamaño actual, terriblemente ineficiente en combustible", sugiere que controle el fuego y libere un mayor valor nutricional la cocina era la adaptación clave que separaba al Homo de los australopitecos que dormían en los árboles.
Un famoso ejemplo de Homo erectus es el Hombre de Pekín; otros se encontraron en Asia (especialmente en Indonesia), África y Europa. Muchos paleoantropólogos ahora usan el término Homo ergaster para las formas no asiáticas de este grupo, y reserva Homo erectus solo para aquellos fósiles que se encuentran en Asia y cumplen con ciertos requisitos esqueletales y dentales que difieren ligeramente de H. ergaster .
H. cepranensis y H. antecessor
Estos se proponen como especies que pueden ser intermedias entre H. erectus y H. heidelbergensis .
- H. antecessor se conoce de fósiles de España e Inglaterra con fecha de 1.2 Ma-500 ka.
- H. cepranensis se refiere a un único casquete craneal de Italia, que se estima tiene unos 800,000 años de antigüedad.
H. heidelbergensis
H. heidelbergensis ("Hombre de Heidelberg") vivió entre 800,000 y 300,000 años atrás. También propuesto como Homo sapiens heidelbergensis o Homo sapiens paleohungaricus .
H. rhodesiensis y el cráneo de Gawis
- H. rhodesiensis , estimado en 300,000-125,000 años. La mayoría de los investigadores actuales ubican al Hombre de Rhodesia dentro del grupo de Homo heidelbergensis , aunque se han propuesto otras designaciones como Homo sapiens arcaico y Homo sapiens rhodesiensis .
- En febrero de 2006, se encontró un fósil, el cráneo de Gawis, que podría ser una especie intermedia entre H. erectus y H. sapiens o uno de los muchos puntos muertos evolutivos. Se cree que el cráneo de Gawis, Etiopía, tiene entre 500,000 y 250,000 años. Solo se conocen los detalles resumidos, y los buscadores aún no han publicado un estudio revisado por pares. Las características faciales del hombre de Gawis sugieren que es una especie intermedia o un ejemplo de una hembra de "hombre Bodo".
Neanderthal y Denisovan

Homo neanderthalensis , alternativamente designado como Homo sapiens neanderthalensis, vivió en Europa y Asia desde hace 400,000 hasta hace unos 28,000 años. Hay una serie de claras diferencias anatómicas entre los humanos anatómicamente modernos (AMH) y las poblaciones de Neanderthal. Muchos de estos se relacionan con la adaptación superior a los ambientes fríos que poseen las poblaciones de Neanderthal. Su relación de superficie a volumen es una versión extrema de la encontrada entre las poblaciones de Inuit, lo que indica que estaban menos inclinadas a perder calor corporal que AMH. Desde Endocasts cerebrales, los neandertales también tenían cerebros significativamente más grandes. Esto parece indicar que la superioridad intelectual de las poblaciones de AMH puede ser cuestionable. Una investigación más reciente de Eiluned Pearce, Chris Stringer, RIM Dunbar, sin embargo, ha mostrado diferencias importantes en la arquitectura del cerebro. Por ejemplo, tanto en el tamaño de la cámara orbital como en el tamaño del lóbulo occipital, el tamaño más grande sugiere que el Neanderthal tenía una mejor agudeza visual que los humanos modernos. Esto daría una visión superior en las condiciones de luz inferiores que se encuentran en Europa Glacial. También parece que la mayor masa corporal de los neandertales tenía una masa cerebral correspondientemente más grande requerida para el cuidado y control del cuerpo.
Las poblaciones de Neanderthal parecen haber sido físicamente superiores a las poblaciones de AMH. Estas diferencias pueden haber sido suficientes para dar a las poblaciones de Neanderthal una superioridad ambiental a las poblaciones de AMH de 75,000 a 45,000 años BP. Con estas diferencias, los cerebros de Neanderthal muestran que había un área más pequeña disponible para el funcionamiento social. Trazar el tamaño del grupo posible a partir del volumen endocrainial sugiere que las poblaciones de AMH (menos el tamaño del lóbulo occipital) tenían un número Dunbar de 144 posibles relaciones. Las poblaciones de neandertales parecen haberse limitado a aproximadamente 120 individuos. Esto se vería en un mayor número de parejas posibles para humanos AMH, con un mayor riesgo de endogamia entre las poblaciones de Neanderthal. También sugiere que los humanos tenían áreas de captación comerciales más grandes que los neandertales (confirmado en la distribución de herramientas de piedra).

La evidencia anterior de la secuenciación del ADN mitocondrial sugirió que no se produjo un flujo significativo de genes entre H. neanderthalensis y H. sapiens , y que las dos eran especies separadas que compartían un ancestro común hace unos 660,000 años. Sin embargo, una secuenciación del genoma del Neandertal en 2010 indica que los neandertales, efectivamente, se entrecruzan con los humanos anatómicamente modernos circa Hace 45,000 a 80,000 años (en el tiempo aproximado en que los humanos modernos emigraron de África, pero antes se dispersaron en Europa, Asia y otros lugares). La secuenciación genética de un esqueleto humano de 40,000 años de Rumania mostró que el 11% de su genoma era Neandertal, y se estimó que el individuo tenía un ancestro Neanderthal 4-6 generaciones anteriores, además de una contribución de un mestizaje previo en el Medio Este. Aunque esta población rumana entrecruzada parece no haber sido ancestral para los humanos modernos, el hallazgo indica que el entrecruzamiento ocurrió repetidamente.
Casi todos los humanos no africanos modernos tienen 1% a 4% de su ADN derivado del ADN de Neanderthal, y este hallazgo es consistente con estudios recientes que indican que la divergencia de algunos alelos humanos data de un Ma, aunque la interpretación de estos estudios ha sido cuestionado Los neandertales y el Homo sapiens podrían haber coexistido en Europa durante 10.000 años, durante los cuales las poblaciones humanas explotaron muchísimo más que los neandertales, posiblemente superándolas por pura fuerza numérica.
En 2008, los arqueólogos que trabajan en el sitio de Denisova Cave en las montañas de Altai de Siberia descubrieron un pequeño fragmento de hueso del quinto dedo de un miembro juvenil de Denisovans. Los artefactos, incluyendo un brazalete, excavado en la cueva en el mismo nivel, fueron cargados con carbono a alrededor de 40,000 BP. Como el ADN había sobrevivido en el fragmento fósil debido al clima frío de la Cueva de Denisova, tanto ADN mitocondrial como ADN nuclear se secuenciaron.
Mientras que el punto de divergencia del ADNmt fue inesperadamente profundo en el tiempo, la secuencia genómica completa sugirió que los denisovanos pertenecían al mismo linaje que los neandertales, con los dos divergentes poco después de que su línea se separó del linaje que dio origen a los humanos modernos. Se sabe que los humanos modernos se han superpuesto con los neandertales en Europa y el Cercano Oriente durante posiblemente más de 40,000 años, y el descubrimiento plantea la posibilidad de que los neandertales, los denisovanos y los humanos modernos hayan coexistido y se hayan cruzado. La existencia de esta rama distante crea una imagen mucho más compleja de la humanidad durante el Pleistocenethan tardío que se pensaba anteriormente. También se ha encontrado evidencia de que tanto como el 6% del ADN de algunos melanesios modernos se derivan de Denisovans, lo que indica un mero cruzamiento en el sudeste asiático.
Los alelos que se cree que se originaron en Neanderthals y Denisovans se han identificado en varios loci genéticos en los genomas de humanos modernos fuera de África. Los haplotipos HLA de Denisovans y Neanderthal representan más de la mitad de los alelos HLA de los eurasiáticos modernos, lo que indica una fuerte selección positiva para estos alelos introgresados. Corinne Simoneti, de la Universidad de Vanderbilt, en Nashville, y su equipo han encontrado en registros médicos de 28,000 personas de ascendencia europea que la presencia de segmentos de ADN de Neanderthal puede asociarse con una probabilidad de sufrir depresión con mayor frecuencia.
El flujo de genes de las poblaciones de Neanderthal a los humanos modernos no fue de una sola dirección. Sergi Castellano del Instituto Max Planck de Antropología Evolutiva en Leipzig, Alemania, en 2016 informó que aunque los genomas de Denisovan y Neanderthal están más relacionados entre sí que lo que son para nosotros, los genomas de Neandertal de Siberia muestran similitud con el conjunto de genes humanos modernos, más así que a las poblaciones europeas de Neanderthal. La evidencia sugiere que las poblaciones de Neanderthal se cruzaron con humanos modernos posiblemente hace 100.000 años, probablemente en algún lugar del Cercano Oriente.
Los estudios de un niño de Neanderthal en Gibraltar muestran a partir del desarrollo del cerebro y la erupción de los dientes que los niños de Neanderthal pueden haber madurado más rápidamente que en el caso del Homo sapiens.
H. floresiensis

H. floresiensis , que vivió desde aproximadamente 190,000 a 50,000 años antes del presente, ha sido apodado hobbit por su pequeño tamaño, posiblemente como resultado del enanismo insular. H. floresiensis intriga tanto por su tamaño como por su edad, siendo un ejemplo de una especie reciente del género Homo que exhibe rasgos derivados que no se comparten con los humanos modernos. En otras palabras, H. floresiensis comparte un ancestro común con los humanos modernos, pero se separó del linaje humano moderno y siguió un camino evolutivo distinto. El hallazgo principal fue un esqueleto que se cree que es una mujer de unos 30 años de edad. Encontrado en 2003, se ha fechado aproximadamente 18,000 años viejo. Se estimó que la mujer viviente tenía un metro de altura, con un volumen cerebral de solo 380 cm (considerado pequeño para un chimpancé y menos de un tercio del promedio de H. sapiens de 1400 cm).
Sin embargo, hay un debate en curso sobre si H. floresiensis es una especie separada. Algunos científicos sostienen que H. floresiensis era un moderno H. sapiens con enanismo patológico. Esta hipótesis es apoyada en parte, porque algunos humanos modernos que viven en Flores, la isla de Indonesia donde se encontró el esqueleto, son pigmeos. Esto, junto con el enanismo patológico, podría haber resultado en un humano significativamente diminuto. El otro ataque importante a H. floresiensis como una especie separada es que se encontró con herramientas asociadas solo con H. sapiens .
La hipótesis del enanismo patológico, sin embargo, no explica las características anatómicas adicionales que son diferentes a las de los humanos modernos (enfermos o no) pero muy similares a los de los miembros antiguos de nuestro género. Además de las características craneales, estas características incluyen la forma de los huesos en la muñeca, el antebrazo, el hombro, las rodillas y los pies. Además, esta hipótesis no explica el hallazgo de múltiples ejemplos de individuos con estas mismas características, lo que indica que eran comunes a una población grande y no se limitaban a un individuo.
H. sapiens
H. sapiens (el adjetivo sapiens es en latín "sabio" o "inteligente") surgió hace unos 300.000 años, probablemente derivado de Homo heidelbergensis . Entre 400,000 años atrás y el segundo período interglacial en el Pleistoceno medio, hace unos 250,000 años, la tendencia en la expansión del volumen intracraneal y la elaboración de tecnologías de herramientas de piedra se desarrollaron, proporcionando evidencia de una transición de H. erectus a H. sapiens . La evidencia directa sugiere que hubo una migración de H. erectus fuera de África, luego una especiación adicional de H. sapiens de H. erectus en África. Una migración posterior (tanto dentro como fuera de África) eventualmente reemplazó al anterior H. erectus disperso . Esta teoría de la migración y el origen suele denominarse la "hipótesis reciente de origen único" o "fuera de África". H. sapiens se cruzó con humanos arcaicos tanto en África como en Eurasia, en Eurasia, especialmente con los neandertales y los denisovanos.
La teoría de la catástrofe de Toba, que postula un cuello de botella poblacional para H. sapiens hace unos 70,000 años, fue controvertida desde su primera propuesta en la década de 1990 y para la de 2010 tuvo muy poco apoyo.
La variabilidad genética humana distintiva ha surgido como resultado del efecto fundador, por la mezcla arcaica y por las recientes presiones evolutivas.
Uso de herramientas




El uso de herramientas se ha interpretado como un signo de inteligencia, y se ha teorizado que el uso de herramientas puede haber estimulado ciertos aspectos de la evolución humana, especialmente la expansión continua del cerebro humano. La paleontología todavía tiene que explicar la expansión de este órgano durante millones de años a pesar de ser extremadamente exigente en términos de consumo de energía. El cerebro de un humano moderno consume alrededor de 13 vatios (260 kilocalorías por día), una quinta parte del consumo de energía en reposo del cuerpo. El aumento en el uso de herramientas permitiría buscar productos cárnicos ricos en energía, y permitiría procesar productos vegetales más ricos en energía. Los investigadores han sugerido que los primeros homínidos estaban bajo presión evolutiva para aumentar su capacidad de crear y usar herramientas.
Precisamente cuando los primeros humanos comenzaron a usar herramientas es difícil de determinar, porque mientras más primitivas sean estas herramientas (por ejemplo, piedras de bordes afilados), más difícil es decidir si son objetos naturales o artefactos humanos. Existe alguna evidencia de que los australopitecinos (4 Ma) pueden haber usado huesos rotos como herramientas, pero esto es debatido.
Cabe señalar que muchas especies fabrican y utilizan herramientas, pero es el género humano el que domina las áreas de fabricación y uso de herramientas más complejas. Las herramientas más antiguas que se conocen son las herramientas de piedra Oldowan de Etiopía, de 2.5-2.6 millones de años. Se encontró un Homo fósil cerca de algunas herramientas de Oldowan, y su edad se observó con 2,3 millones de años, lo que sugiere que tal vez el Homo las especies sí crearon y usaron estas herramientas. Es una posibilidad, pero aún no representa una evidencia sólida. El tercer proceso estiloide metacarpiano permite que el hueso de la mano se fije en los huesos de la muñeca, lo que permite aplicar una mayor cantidad de presión en la muñeca y la mano desde el pulgar y los dedos. Le permite a los humanos la destreza y la fuerza para hacer y usar herramientas complejas. Esta característica anatómica única separa a los humanos de los simios y otros primates no humanos, y no se ve en fósiles humanos de más de 1,8 millones de años.
Bernard Wood notó que Paranthropus coexistió con las primeras especies Homo en el área del "Complejo Industrial Oldowan" durante aproximadamente el mismo lapso de tiempo. Aunque no existe evidencia directa que identifique a Paranthropus como los fabricantes de herramientas, su anatomía se presta a la evidencia indirecta de sus capacidades en esta área. La mayoría de los paleoantropólogos coinciden en que las primeras especies de Homo fueron responsables de la mayoría de las herramientas de Oldowan encontradas. Argumentan que cuando la mayoría de las herramientas de Oldowan se encontraron en asociación con fósiles humanos, el Homo siempre estaba presente, pero Paranthropus no.
En 1994, Randall Susman utilizó la anatomía de los pulgares oponibles como la base de su argumento de que tanto la especie Homo como la de Paranthropus eran fabricantes de herramientas. Comparó huesos y músculos de pulgares humanos y de chimpancé, descubriendo que los humanos tienen 3 músculos que carecen de chimpancés. Los humanos también tienen metacarpianos más gruesos con cabezas más anchas, lo que permite un agarre más preciso del que la mano del chimpancé puede realizar. Susman postuló que la anatomía moderna del pulgar oponible humano es una respuesta evolutiva a los requisitos asociados con las herramientas de fabricación y manipulación y que ambas especies eran de hecho hacedores de herramientas.
Herramientas de piedra
Las herramientas de piedra se atestiguaron por primera vez hace unos 2,6 millones de años, cuando H. habilis en el este de África utilizó las llamadas herramientas de guijarros, helicópteros hechos de guijarros redondos que se habían partido por simples golpes. Esto marca el comienzo del Paleolítico, o Vieja Edad de Piedra; se considera que su fin es el final de la última Edad de Hielo, hace unos 10.000 años. El Paleolítico se subdivide en el Paleolítico Inferior (Edad de Piedra Inicial), terminando hace aproximadamente 350,000-300,000 años, el Paleolítico Medio (Edad de Piedra Medio), hasta 50,000-30,000 años atrás, y el Paleolítico Superior, (Edad de Piedra Tardía), 50,000 -10,000 años atrás.
Los arqueólogos que trabajan en el Gran Valle del Rift en Kenia afirman haber descubierto las herramientas de piedra más antiguas que se conocen en el mundo. Con una antigüedad de alrededor de 3,3 millones de años, los implementos son unos 700,000 años más antiguos que las herramientas de piedra de Etiopía que anteriormente tenían esta distinción.
El período de hace 700,000-300,000 años también se conoce como Acheulean, cuando H. ergaster (o erectus ) hizo grandes hachas de piedra con cuarcitas y pedernal, al principio bastante ásperas (Early Acheulian), más tarde "retocadas" por adicional, golpes más sutiles a los lados de los copos. Después de 350,000 BP, se desarrolló la técnica más refinada de Levallois, una serie de golpes consecutivos, mediante los cuales se hicieron raspadores, rebanadores ("racloirs"), agujas y agujas aplanadas. Finalmente, después de aproximadamente 50,000 BP, los neandertales y los inmigrantes Cro-Magnons (cuchillos, cuchillas, espumaderas) fabricaron herramientas de pedernal más refinadas y especializadas. En este período, también comenzaron a fabricar herramientas con hueso.
Transición a la modernidad conductual
Hasta hace unos 50,000-40,000 años, el uso de herramientas de piedra parece haber progresado paso a paso. Cada fase ( H. habilis , H. ergaster , H. neanderthalensis ) comenzó en un nivel más alto que el anterior, pero después de cada fase comenzó, el desarrollo posterior fue lento. Actualmente, los paleoantropólogos debaten si estas especies Homo poseían algunos o muchos de los rasgos culturales y de comportamiento asociados con los humanos modernos, como el lenguaje, el pensamiento simbólico complejo, la creatividad tecnológica, etc. Parece que eran culturalmente conservadores manteniendo tecnologías simples y patrones de alimentación durante mucho tiempo períodos.
Alrededor de 50,000 BP, la cultura humana moderna comenzó a evolucionar más rápidamente. La transición a la modernidad del comportamiento ha sido caracterizada por la mayoría como un "Gran Salto Adelante" de Eurasia, o como la "Revolución Paleolítica Superior", debido a la aparición repentina de signos distintivos de la conducta moderna y la caza mayor en el registro arqueológico. Algunos otros estudiosos consideran que la transición ha sido más gradual, señalando que algunas características ya habían aparecido entre el Homo sapiens africano arcaico desde hace 200.000 años. La evidencia reciente sugiere que la población aborigen australiana se separó de la población africana hace 75,000 años, y que hicieron un viaje por mar de hasta 160 km hace 60,000 años, lo que puede disminuir la evidencia de la Revolución del Paleolítico Superior.
Los humanos modernos comenzaron a enterrar a sus muertos, usando pieles de animales para confeccionar ropas, cazando con técnicas más sofisticadas (como el uso de pozos de trampas o alejando a los animales de los acantilados) y participando en la pintura de cuevas. A medida que avanzó la cultura humana, diferentes poblaciones de humanos introdujeron la novedad a las tecnologías existentes: artefactos como anzuelos, botones y agujas de hueso muestran signos de variación entre diferentes poblaciones de humanos, algo que no se había visto en las culturas humanas antes de 50,000 BP. Típicamente, las poblaciones de H. neanderthalensis no varían en sus tecnologías, aunque se ha encontrado que los ensamblajes Chatelperronianos son innovaciones neandertales producidas como resultado de la exposición a las tecnologías Homo sapiens Aurignacian.
Entre los ejemplos concretos del comportamiento humano moderno, los antropólogos incluyen la especialización de herramientas, el uso de joyas e imágenes (como dibujos de cuevas), organización del espacio vital, rituales (por ejemplo, enterramientos con dones graves), técnicas de caza especializadas, exploración de menos hospitalarios áreas geográficas y redes de bartertrade. Continúa el debate sobre si una "revolución" llevó a los humanos modernos ("el big bang de la conciencia humana"), o si la evolución fue más "gradual".
Evolución humana reciente y actual
La evolución ha continuado en poblaciones humanas anatómicamente modernas, que se ven afectadas tanto por la selección natural como por la deriva genética. Aunque la presión de selección sobre algunos rasgos, como la resistencia a la viruela, ha disminuido en la vida humana moderna, los humanos todavía están sometidos a selección natural para muchos otros rasgos. Algunos de ellos se deben a presiones ambientales específicas, mientras que otros están relacionados con cambios en el estilo de vida desde el desarrollo de la agricultura (hace 10.000 años), la civilización urbana (5,000) y la industrialización (hace 250 años). Se ha argumentado que la evolución humana se ha acelerado desde el desarrollo de la agricultura hace 10.000 años y la civilización hace unos 5.000 años, lo que resulta, según se afirma, en diferencias genéticas sustanciales entre las diferentes poblaciones humanas actuales.
Particularmente conspicua es la variación en las características superficiales, como el pelo Afro-textured, o la evolución reciente de la piel clara y el pelo rubio en algunas poblaciones, que se atribuyen a las diferencias en el clima. Presiones selectivas particularmente fuertes han resultado en la adaptación a gran altitud en humanos, con diferentes en diferentes poblaciones aisladas. Los estudios de la base genética muestran que algunos se desarrollaron muy recientemente, y los tibetanos evolucionaron durante 3.000 años para tener altas proporciones de un alelo de EPAS1 que es adaptable a grandes altitudes.
Otra evolución se relaciona con enfermedades endémicas: la presencia de malaria seleccionada para el rasgo drepanocítico (la forma heterocigótica del gen drepanocítico), mientras que la ausencia de malaria y los efectos sobre la salud de la anemia drepanocítica se eligen en contra de este rasgo. Por ejemplo, la población en riesgo de la enfermedad debilitante grave kuru tiene una sobrerrepresentación significativa de una variante inmune del gen de la proteína prión G127V frente a los alelos no inmunes. La frecuencia de esta variante genética se debe a la supervivencia de las personas inmunes.
La evolución humana reciente relacionada con la agricultura incluye resistencia genética a enfermedades infecciosas que ha aparecido en poblaciones humanas al cruzar la barrera de especies de animales domésticos, así como cambios en el metabolismo debido a cambios en la dieta, como la persistencia de lactasa.
En la época contemporánea, desde la industrialización, se han observado algunas tendencias: por ejemplo, la menopausia está evolucionando para ocurrir más tarde. Otras tendencias informadas parecen incluir el alargamiento del período reproductivo humano y la reducción en los niveles de colesterol, glucosa en sangre y presión arterial en algunas poblaciones.
Lista de especies
Esta lista está en orden cronológico en la tabla por género . Algunos nombres de especies / subespecies están bien establecidos, y algunos están menos establecidos, especialmente en el género Homo . Por favor vea los artículos para más información.
| Sahelanthropus | Homo (humano) |
|---|---|
| S. tchadensis | H. gautengensis |
| Orrorin | H. habilis |
| O. tugenensis | H. rudolfensis |
| Ardipithecus | H. floresiensis |
| A. kadabba | H. ergaster |
| A. ramidus | H. erectus |
| Australopithecus | • H. e. georgicus |
| A. anamensis | H. cepranensis |
| A. afarensis | H. antecessor |
| A. bahrelghazali | H. heidelbergensis |
| A. africanus | H. rhodesiensis |
| A. garhi | H. naledi |
| A. sediba | H. helmei |
| Kenyanthropus | H. neanderthalensis |
| K. platyops | H. sapiens |
| Paranthropus | • H. s. Idaltu |
| P. aethiopicus | • H. s. sapiens (temprano) |
| P. boisei | • H. s. sapiens (moderno) |
| P. robustus |