Eucariota
Definición
| Eukaryotes Rango temporal: Orosirian - Presente 1850-0Ma Had'n Archean Proterozoico Pha. | |
|---|---|
 | |
| Eucariotas y algunos ejemplos de su diversidad - en el sentido de las agujas del reloj desde arriba a la izquierda: abeja albañil roja, Boletus edulis , chimpancé común, Isotricha intestinalis , ranúnculo persa y Volvox carteri | |
| clasificación cientifica | |
| Dominio: | Eukaryota (Chatton, 1925) Whittaker y Margulis, 1978 |
| Supergrupos y reinos | |
Los organismos eucariotas que no pueden clasificarse en los reinos Plantae, Animalia o Hongos a veces se agrupan en el reino Protista . | |
Los eucariotas pueden reproducirse asexualmente a través de mitosis y sexualmente a través de la meiosis y la fusión de gametos. En la mitosis, una célula se divide para producir dos células genéticamente idénticas. En la meiosis, la replicación del ADN es seguida por dos rondas de división celular para producir cuatro células hijas haploides. Estos actúan como células sexuales (gametos). Cada gameto tiene solo un conjunto de cromosomas, cada uno una combinación única del par correspondiente de cromosomas parentales resultante de la recombinación genética durante la meiosis.
El dominio Eukaryota parece ser monofilético y constituye uno de los dominios de la vida en el sistema de tres dominios. Los otros dos dominios, Bacteria y Archaea, son procariotas y no tienen ninguna de las características anteriores. Los eucariotas representan una pequeña minoría de todos los seres vivos. Sin embargo, debido a su tamaño en general mucho más grande, se estima que su biomasa mundial colectiva es aproximadamente igual a la de los procariotas. Los eucariotas evolucionaron aproximadamente hace 1.6-2.1 mil millones de años, durante el eón Proterozoico.
Historia

En 1905 y 1910, el biólogo ruso Konstantin Mereschkowski (1855-1921) argumentó que los plástidos eran cianobacterias reducidas en una simbiosis con un hospedador no fotosintético (heterótrofo) que se formaba por simbiosis entre un hospedador similar a una ameba y una célula similar a una bacteria que formó el núcleo. Las plantas habían heredado la fotosíntesis de las cianobacterias.
El concepto de eucariota ha sido atribuido al biólogo francés Edouard Chatton (1883-1947). Los términos procariota y eucariota fueron reintroducidos de manera más definitiva por el microbiólogo canadiense Roger Stanier y el microbiólogo holandés-estadounidense CB van Niel en 1962. En su obra Titres et Travaux Scientifiques de 1938 , Chatton propuso los dos términos, llamando a las bacterias procariotas y organismos con núcleos en sus células eucariotas. Sin embargo, mencionó esto en solo un párrafo, y la idea fue efectivamente ignorada hasta que la declaración de Chatton fue redescubierta por Stanier y van Niel.
En 1967, Lynn Margulis proporcionó pruebas microbiológicas de la endosimbiosis como origen de los cloroplastos y las mitocondrias en células eucariotas en su artículo sobre el origen de las células mitoscentes. En la década de 1970, Carl Woese exploró la filogenia microbiana, estudiando las variaciones en el ARN ribosómico 16S. Esto ayudó a descubrir el origen de los eucariotas y la simbiosis de dos importantes orgánulos eucariotas, mitocondrias y cloroplastos. En 1977, Woese y George Fox introdujeron una "tercera forma de vida", a la que llamaron Archaebacteria; en 1990, Woese, Otto Kandler y Mark L. Wheeler lo llamaron Archaea.
En 1979, GW Gould y GJ Dring sugirieron que el núcleo de la célula eucariótica provenía de la capacidad de las bacterias Gram-positivas para formar endosporas. En 1987 y documentos posteriores, Thomas Cavalier-Smith propuso en cambio que las membranas del núcleo y del retículo endoplasmático se formaron primero al inflar la membrana plasmática de un procariota. En la década de 1990, varios otros biólogos propusieron orígenes endosimbióticos para el núcleo, reviviendo efectivamente la teoría de Mereschkowsky.
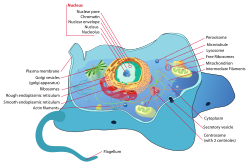
Características de la celda
Las células eucariotas son típicamente mucho más grandes que las de los procariotas que tienen un volumen de alrededor de 10.000 veces mayor que la célula procariota. Tienen una variedad de estructuras internas unidas a la membrana, llamadas organelas, y un citoesqueleto compuesto de microtúbulos, microfilamentos y filamentos intermedios, que desempeñan un papel importante en la definición de la organización y la forma de la célula. El ADN eucariótico se divide en varios haces lineales llamados cromosomas, que están separados por un huso microtubular durante la división nuclear.
Membrana interna

Las células eucariotas incluyen una variedad de estructuras unidas a la membrana, denominadas colectivamente sistema de endomembrana. Se pueden formar compartimentos simples, llamados vesículas y vacuolas, brotando de otras membranas. Muchas células ingieren alimentos y otros materiales a través de un proceso de endocitosis, donde la membrana externa se invagina y luego se separa para formar una vesícula. Es probable que la mayoría de los otros orgánulos unidos a la membrana se deriven finalmente de tales vesículas. Alternativamente, algunos productos producidos por la célula pueden dejarse en una vesícula a través de la exocitosis.
El núcleo está rodeado por una doble membrana (comúnmente conocida como membrana nuclear o envoltura nuclear), con poros que permiten que el material se mueva hacia adentro y hacia afuera. Varias extensiones en forma de tubo y lámina de la membrana nuclear forman el retículo endoplásmico, que está involucrado en el transporte y la maduración de las proteínas. Incluye el retículo endoplasmático rugoso donde se unen los ribosomas para sintetizar proteínas, que ingresan al espacio interior o luz. Posteriormente, generalmente entran vesículas, que brotan del retículo endoplásmico liso. En la mayoría de los eucariotas, estas vesículas portadoras de proteínas se liberan y se modifican adicionalmente en pilas de vesículas aplanadas (cisterna), el aparato de Golgi.
Las vesículas pueden estar especializadas para diversos fines. Por ejemplo, los lisosomas contienen enzimas digestivas que descomponen la mayoría de las biomoléculas en el citoplasma. Los peroxisomas se usan para descomponer el peróxido, que de lo contrario es tóxico. Muchos protozoos tienen vacuolas contráctiles, que recogen y expulsan el exceso de agua, y extrusomas, que expulsan el material utilizado para desviar a los depredadores o capturar presas. En las plantas superiores, la mayor parte del volumen de una celda es absorbida por una vacuola central, que en su mayoría contiene agua y principalmente mantiene su presión osmótica.
Mitocondrias y plastos

Las mitocondrias son organelos que se encuentran en todos menos en un eucarionte. Las mitocondrias proporcionan energía a la célula eucariota mediante la conversión de azúcares en ATP. Tienen dos membranas circundantes, cada una una bi-capa de fosfolípidos; el interior del cual se pliega en invaginaciones llamadas crestas donde tiene lugar la respiración aeróbica.
Las mitocondrias contienen su propio ADN, que tiene similitudes estructurales estrechas con el ADN bacteriano, y que codifica los genes ARNr y ARNt que producen ARN que tiene una estructura más cercana al ARN bacteriano que al ARN eucariota. En general, se considera que se desarrollaron a partir de procariotas endosimbióticos, probablemente proteobacterias.
Algunos eucariotas, como los metamonads como Giardia y Trichomonas , y el amebozoan Pelomyxa, parecen carecer de mitocondrias, pero se ha encontrado que todos ellos contienen orgánulos derivados de la mitocondria, como hidrogenosomas y mitosomas, y por lo tanto han perdido sus mitocondrias de forma secundaria. Obtienen energía por acción enzimática sobre los nutrientes absorbidos del medio ambiente. El metamonad Monocercomonoides también ha adquirido, mediante transferencia génica lateral, un sistema de movilización de azufre citosólico que proporciona los agregados de hierro y azufre necesarios para la síntesis de proteínas. La ruta del clúster de hierro-azufre mitocondrial normal se ha perdido de forma secundaria.
Las plantas y varios grupos de algas también tienen plastidios. Los plástidos también tienen su propio ADN y se desarrollan a partir de endosimbiontes, en este caso cianobacterias. Por lo general, toman la forma de cloroplastos que, como las cianobacterias, contienen clorofila y producen compuestos orgánicos (como la glucosa) a través de la fotosíntesis. Otros están involucrados en el almacenamiento de alimentos. Aunque los plástidos probablemente tengan un único origen, no todos los grupos que contienen plastos están estrechamente relacionados. En cambio, algunos eucariotas los han obtenido de otros mediante endosimbiosis secundaria o ingestión. La captura y secuestro de células fotosintéticas y cloroplastos ocurre en muchos tipos de organismos eucarióticos modernos y se conoce como cleptoplastia.
Los orígenes endosimbióticos también se han propuesto para el núcleo y para el flagelo eucariótico.
Estructuras del citoesqueleto

Muchos eucariotes tienen proyecciones citoplásmicas largas y delgadas y delgadas, llamadas flagelos, o estructuras similares llamadas cilios. Los flagelos y los cilios a veces se denominan undulipodia y están involucrados de diversas maneras en el movimiento, la alimentación y la sensación. Están compuestos principalmente de tubulina. Estos son completamente distintos de los flagelos procarióticos. Se apoyan en un manojo de microtúbulos que surgen de un centríolo, característicamente dispuestos como nueve dobletes que rodean dos singletes. Flagella también puede tener pelos o mastigonemas y escamas que conectan membranas y varillas internas. Su interior es continuo con el citoplasma de la célula.
Las estructuras microfilamentarias compuestas de proteínas de unión a actina y actina, por ejemplo, a-actinina, fimbrina, filamina, están presentes también en capas corticales submembranas y haces. Las proteínas motoras de los microtúbulos, por ejemplo, dineína o quinesina y actina, por ejemplo, miosinas, proporcionan un carácter dinámico de la red.
Los centríolos a menudo están presentes incluso en células y grupos que no tienen flagelos, pero las coníferas y las plantas con flores no tienen ninguno. Generalmente ocurren en grupos que dan lugar a diversas raíces microtubulares. Estos forman un componente principal de la estructura del citoesqueleto, y a menudo se ensamblan a lo largo de varias divisiones celulares, con un flagelo retenido del padre y el otro derivado de él. Centrioles producen el huso durante la división nuclear.
La importancia de las estructuras del citoesqueleto se subraya en la determinación de la forma de las células, así como también en que son componentes esenciales de las respuestas migratorias como la quimiotaxis y la quimiocinesis. Algunos protistas tienen otros organelos soportados por microtúbulos. Estos incluyen la radiolaria y heliozoa, que producen axopodia utilizado en flotación o para capturar presas, y los haptófitos, que tienen un organelo peculiar tipo flagelo llamado el haptonema.
Pared celular
Las células de plantas y algas, hongos y la mayoría de los cromalveolados tienen una pared celular, una capa fuera de la membrana celular, que proporciona a la célula soporte estructural, protección y un mecanismo de filtración. La pared celular también evita la sobreexpansión cuando el agua ingresa a la celda.
Los principales polisacáridos que componen la pared celular primaria de las plantas terrestres son celulosa, hemicelulosa y pectina. Las microfibrillas de celulosa se unen por medio de ataduras hemicelulósicas para formar la red de celulosa-hemicelulosa, que está incorporada en la matriz de pectina. La hemicelulosa más común en la pared celular primaria es el xiloglucano.
Diferencias entre las células eucariotas
Hay muchos tipos diferentes de células eucariotas, aunque los animales y las plantas son los eucariotas más familiares, y por lo tanto proporcionan un excelente punto de partida para comprender la estructura eucariota. Sin embargo, los hongos y muchos protistas tienen algunas diferencias sustanciales.
Célula animal

Todos los animales son eucariotas. Las células animales son distintas de las de otros eucariotas, sobre todo las plantas, ya que carecen de paredes celulares y cloroplastos y tienen vacuolas más pequeñas. Debido a la falta de una pared celular, las células animales pueden adoptar una variedad de formas. Una célula fagocítica puede englobar otras estructuras.
Célula vegetal
Las células vegetales son bastante diferentes de las células de los otros organismos eucariotas. Sus características distintivas son:
- Una gran vacuola central (encerrada por una membrana, el tonoplasto), que mantiene la turgencia de la célula y controla el movimiento de moléculas entre el citosol y la savia
- Una pared celular primaria que contiene celulosa, hemicelulosa y pectina, depositada por el protoplasto en el exterior de la membrana celular; esto contrasta con las paredes celulares de los hongos, que contienen quitina, y las envolturas celulares de los procariotas, en los que los peptidoglicanos son las principales moléculas estructurales
- Los plasmodesmos, poros en la pared celular que unen las células adyacentes y permiten que las células vegetales se comuniquen con las células adyacentes. Los animales tienen un sistema diferente y funcionalmente análogo de uniones gap entre células adyacentes.
- Plastidios, especialmente cloroplastos que contienen clorofila, el pigmento que da a las plantas su color verde y les permite realizar la fotosíntesis
- Las briófitas y las plantas vasculares sin semillas solo tienen flagelos y centriolos en las células espermáticas. El esperma de las cícadas y el Ginkgo son células grandes y complejas que nadan con cientos o miles de flagelos.
- Las coníferas (Pinophyta) y las plantas con flores (angiospermas) carecen de los flagelos y centriolos que están presentes en las células animales.
Célula fúngica

Las células de los hongos son más similares a las células animales, con las siguientes excepciones:
- Una pared celular que contiene quitina
- Menos definición entre celdas; las hifas de los hongos superiores tienen particiones porosas llamadas septas, que permiten el paso del citoplasma, los orgánulos y, a veces, los núcleos. Los hongos primitivos tienen pocos o ningún septa, por lo que cada organismo es esencialmente una supercélula gigante multinucleada; estos hongos se describen como cenocíticos.
- Solo los hongos más primitivos, los chitridios, tienen flagelos.
Otras células eucariotas
Algunos grupos de eucariotas tienen organelos únicos, como los cyanelles (cloroplastos inusuales) de los glauofitas, el háptonema de las haptófitas o los expulsadosomas de las criptomonas. Otras estructuras, como los pseudópodos, se encuentran en diversos grupos eucariotas en diferentes formas, como los amebozoos lobulares o los foraminíferos reticulósicos.
Reproducción

La división celular generalmente se lleva a cabo asexualmente por mitosis, un proceso que permite que cada núcleo hijo reciba una copia de cada cromosoma. La mayoría de los eucariotas también tienen un ciclo de vida que implica la reproducción sexual, alternando entre una fase haploide, donde solo está presente una copia de cada cromosoma en cada célula y una fase diploide, en la que dos copias de cada cromosoma están presentes en cada célula. La fase diploide está formada por la fusión de dos gametos haploides para formar un zigoto, que puede dividirse por mitosis o someterse a una reducción cromosómica por meiosis. Hay una variación considerable en este patrón. Los animales no tienen fase haploide multicelular, pero cada generación de plantas puede consistir en fases pluricelulares haploides y diploides.
Los eucariotas tienen una relación menor de área superficial a volumen que los procariotas, y por lo tanto tienen tasas metabólicas más bajas y tiempos de generación más largos.
La evolución de la reproducción sexual puede ser una característica primordial y fundamental de los eucariotas. Basado en un análisis filogenético, Dacks y Roger propusieron que el sexo facultativo estaba presente en el ancestro común de todos los eucariotas. Un conjunto básico de genes que funcionan en la meiosis está presente tanto en Trichomonas vaginalis como en Giardia intestinalis , dos organismos que anteriormente se creía que eran asexuales. Dado que estas dos especies son descendientes de linajes que divergieron temprano del árbol evolutivo eucariótico, se infirió que los genes meióticos centrales, y por lo tanto el sexo, probablemente estaban presentes en un ancestro común de todos los eucariotas. Especies eucariotas que alguna vez se creyeron asexuales, como protozoos parásitos del género Leishmania, se ha demostrado que tienen un ciclo sexual. Además, la evidencia ahora indica que las amebas, previamente consideradas como asexuales, son antiguamente sexuales y que la mayoría de los grupos asexuales actuales probablemente surgieron recientemente e independientemente.
Clasificación



En la antigüedad, los dos linajes de animales y plantas fueron reconocidos. Se les dio el rango taxonómico de Reino por Linnaeus. Aunque incluyó los hongos con plantas con algunas reservas, más tarde se dio cuenta de que son bastante distintos y justifican un reino separado, cuya composición no estaba del todo clara hasta la década de 1980. Los diversos eucariotas de células individuales se colocaron originalmente con plantas o animales cuando se conocieron. En 1830, el biólogo alemán Georg A. Goldfuss acuñó la palabra protozoos para referirse a organismos como ciliados, y este grupo se expandió hasta abarcar a todos los eucariotas unicelulares, y su propio reino, el Protista, por Ernst Haeckel en 1866 Los eucariotas llegaron a estar compuestos por cuatro reinos:
- Reino Protista
- Reino Plantae
- Hongos del Reino
- Reino Animal
Se entendía que los protistas eran "formas primitivas" y, por lo tanto, un grado evolutivo, unidos por su naturaleza unicelular primitiva. El desenmarañamiento de las divisiones profundas en el árbol de la vida solo comenzó realmente con la secuenciación del ADN, lo que condujo a un sistema de dominios en lugar de reinos, como Carl Woese, líder de nivel superior, unificando todos los reinos eucariotas bajo el dominio eucariota. Al mismo tiempo, el trabajo en el árbol protista se intensificó, y todavía continúa activo hoy. Se han enviado varias clasificaciones alternativas, aunque no hay consenso en el campo.
Los eucariotas son un clado generalmente considerado como hermano de Heimdallarchaeota en la agrupación Asgard en las Archaea. Las agrupaciones basales son Opimoda, Diphoda, Discoba y Loukozoa. La raíz eucariota generalmente se considera que está cerca o incluso en Discoba.
Una clasificación producida en 2005 para la Sociedad Internacional de Protistas, que refleja el consenso de la época, dividió a los eucariotas en seis "supergrupos" supuestamente monofiléticos. Sin embargo, en el mismo año (2005), se expresaron dudas sobre si algunos de estos supergrupos eran monofiléticos, particularmente el Chromalveolata, y una revisión en 2006 señaló la falta de pruebas para varios de los supuestos seis supergrupos. Una clasificación revisada en 2012 reconoce cinco supergrupos.
| Archaeplastida (o Primoplantae) | Plantas terrestres, algas verdes, algas rojas y glaucofitas |
| Supergrupo SAR | Stramenopiles (algas marrones, diatomeas, etc.), Alveolata y Rhizaria (Foraminifera, Radiolaria y varios otros protozoos ameboides). |
| Excavata | Varios protozoos flagelados |
| Amoebozoa | La mayoría de los amoeboides loboseos y mohos de limo |
| Opisthokonta | Animales, hongos, choanoflagelados, etc. |
También hay grupos más pequeños de eucariotas cuya posición es incierta o parece quedar fuera de los grupos principales, en particular, Haptophyta, Cryptophyta, Centrohelida, Telonemia, Picozoa, Apusomonadida, Ancyromonadida, Breviatea y el género Collodictyon . En general, parece que, aunque se ha avanzado, todavía hay incertidumbres muy significativas en la historia evolutiva y la clasificación de eucariotas. Como Roger y Simpson dijeron en 2009 "con el ritmo actual de cambio en nuestra comprensión del árbol eucariota de la vida, debemos proceder con precaución".
En un artículo publicado en Nature Microbiology en abril de 2016, los autores "reforzaron una vez más que la vida que vemos a nuestro alrededor -plantas, animales, humanos y otros supuestos eucariotas- representa un pequeño porcentaje de la biodiversidad del mundo". Clasificaron el eucariota "basado en la herencia de su sistemas de información en lugar de lípidos u otras estructuras celulares ". Jillian F. Banfield de la Universidad de California, Berkeley y sus colegas científicos utilizaron una supercomputadora para generar un diagrama de un nuevo árbol de la vida basado en ADN de 3000 especies, incluidas 2.072 especies conocidas y 1.011 organismos microbianos recientemente informados, cuyo ADN habían reunido de diversos entornos. A medida que la capacidad de secuenciar el ADN se hizo más fácil, Banfield y su equipo pudieron hacer secuencias metagenómicas- "
Filogenia
Los árboles de ARNr construidos durante las décadas de 1980 y 1990 dejaron a la mayoría de los eucariotas en un grupo de "corona" no resuelto (técnicamente no era una verdadera corona), que generalmente se dividía por la forma de las crestas mitocondriales; ver coronas eucariotas. Los pocos grupos que carecen de mitocondrias se ramificaron por separado, por lo que se creía que la ausencia era primitiva; pero esto ahora se considera un artefacto de atracción de rama larga, y se sabe que los perdieron de manera secundaria.
A partir de 2011, existe un acuerdo generalizado de que la Rhizaria pertenece con los Stramenopiles y la Alveolata, en un clado denominado el supergrupo SAR, por lo que Rhizaria no es uno de los principales grupos eucariotas; también que los Amoebozoa y Opisthokonta son monofiléticos y forman un clado, a menudo llamados unikonts. Más allá de esto, no parece haber consenso.
Se ha estimado que puede haber 75 linajes distintos de eucariotas. La mayoría de estos linajes son protistas.
Los tamaños conocidos del genoma eucariota varían desde 8,2 megabases (Mb) en Babesia bovis hasta 112 000-220 050 Mb en el dinoflagelado Prorocentrum micans , lo que demuestra que el genoma del eucariota ancestral ha sufrido variaciones considerables durante su evolución. El último ancestro común de todos los eucariotas se cree que fue un protista fagotrófico con un núcleo, al menos un centriolo y cilio, mitocondrias facultativamente aeróbicas, sexo (meiosis y singamia), un quiste dormido con una pared celular de quitina y / o celulosa y peroxisomas. La endosimbiosis posterior condujo a la propagación de los plástidos en algunos linajes.
Cinco supergrupos
Un árbol global de eucariotas a partir de un consenso de evidencia filogenética (en particular, filogenómica), firmas genómicas raras y características morfológicas se presenta en Adl et al. 2012 y Burki 2014/2016 con Cryptophyta y picozoa surgiendo dentro de la Archaeplastida. Una inclusión similar de Glaucophyta, Cryptista (y también, inusualmente, Haptista) también se ha hecho.
| Eucariotas |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||




En algunos análisis, el grupo Hacrobia (Haptophyta + Cryptophyta) se coloca junto a Archaeplastida, pero en otros está anidado dentro de Archaeplastida. Sin embargo, varios estudios recientes han concluido que Haptophyta y Cryptophyta no forman un grupo monofilético. El primero podría ser un grupo hermano del grupo SAR, el segundo grupo con el Archaeplastida (plantas en el sentido amplio).
La división de los eucariontes en dos clados primarios, bikontes (Archaeplastida + SAR + Excavata) y unikonts (Amoebozoa + Opisthokonta), derivados de un organismo biflagellar ancestral y un organismo ancestral uniflagellar, respectivamente, se había sugerido anteriormente. Un estudio de 2012 produjo una división algo similar, aunque señaló que los términos "unikonts" y "bikonts" no se usaron en el sentido original.
Un conjunto de árboles altamente convergentes y congruentes aparece en Derelle et al (2015), Ren et al (2016), Yang et al (2017) y Cavalier-Smith (2015) incluyendo la información complementaria, lo que resulta en un árbol más conservador y consolidado . Se combina con algunos resultados de Cavalier-Smith para el Opimoda basal. Las principales controversias restantes son la raíz y el posicionamiento exacto de la Rhodophyta y los bikontes Rhizaria, Haptista, Cryptista, Picozoa y Telonemia, muchos de los cuales pueden ser híbridos endosymbyotic eukaryote-eucariotas. Archeaplastida desarrolló los cloroplastos probablemente por endosimbiosis de un ancestro relacionado con una cianobacteria actualmente existente, Gloeomargarita lithophora.
| Eucariotas |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Árbol de Cavalier-Smith
Thomas Cavalier-Smith 2010, 2013, 2014, 2017 y 2018 coloca la raíz del árbol eucariota entre Excavata (con surco de alimentación ventral sostenido por una raíz microtubular) y la Euglenozoa sin estrías y la Chromista monofilética, correlacionada con un solo evento endosymbyótico de capturar un rojo -algas. He et al apoya específicamente el enraizamiento del árbol eucariótico entre un Discoba monofilético (Discicristata + Jakobida) y un clado de Amorphea-Diaphoretickes.
| Eucariotas |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Origen de eucariotas


Fósiles
El origen de la célula eucariota es un hito en la evolución de la vida, ya que los eucariotas incluyen todas las células complejas y casi todos los organismos multicelulares. El tiempo de esta serie de eventos es difícil de determinar; Knoll (2006) sugiere que se desarrollaron hace aproximadamente 1.6-2.100 millones de años. Algunos acritarcos son conocidos hace al menos 1,65 mil millones de años, y la posible alga Grypania se remonta a hace 2,100 millones de años. El hongo fósil similar a Geosiphon, Diskagma, se ha encontrado en paleosuelos de hace 2.200 millones de años.
Se han encontrado estructuras vivas organizadas en las lutitas negras de la Formación Palaeoproterozoica Francevillian B en Gabón, fechadas en 2.100 millones de años. La vida eucariótica podría haber evolucionado en ese momento. Fósiles que están claramente relacionados con grupos modernos comienzan a aparecer hace un estimado de 1.2 mil millones de años, en la forma de un alga roja, aunque trabajos recientes sugieren la existencia de algas filamentosas fosilizado en la cuenca de citas Vindhya remonta quizás a hace 1,6 a 1,7 mil millones de años.
Los biomarcadores sugieren que al menos los eucariotas del tallo surgieron incluso antes. La presencia de esteras en las lutitas australianas indica que los eucariotas estuvieron presentes en estas rocas con una edad de 2.700 millones de años, aunque se sugirió que podrían originarse a partir de la contaminación de las muestras.
Siempre que sus orígenes, eucariotas pueden no haber llegado a ser ecológicamente dominante hasta mucho más tarde; un aumento masivo en la composición de zinc de los sedimentos marinos hace 800 millones de años se ha atribuido al aumento de poblaciones sustanciales de eucariotas, que preferentemente consumen e incorporan zinc en relación con los procariotas.
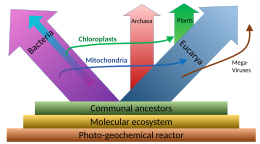
Relación con Archaea
El ADN nuclear y la maquinaria genética de los eucariotas son más similares a Archaea que a las bacterias, lo que lleva a una sugerencia controvertida de que los eucariotas se deben agrupar con Archaea en el clado Neomura. En otros aspectos, como la composición de la membrana, los eucariotas son similares a las bacterias. Se han propuesto tres explicaciones principales para esto:
- Los eucariotas resultaron de la fusión completa de dos o más células, en donde el citoplasma se formó a partir de una eubacteria, y el núcleo de una archaeon, de un virus, o de una pre-célula.
- Los eucariotas se desarrollaron a partir de Archaea, y adquirieron sus características eubacteriales a través de la endosimbiosis de una proto-mitocondria de origen eubacterial.
- Eukaryotes y Archaea se desarrollaron por separado de una eubacteria modificada.
Las propuestas alternativas incluyen:
- La hipótesis del cronocito postula que una célula eucariota primitiva se formó por la endosimbiosis de ambas arqueas y bacterias por un tercer tipo de célula, denominado cronocito.
- El ancestro común universal (UCA) del árbol de la vida actual era un organismo complejo que sobrevivió a un evento de extinción masiva en lugar de una etapa temprana en la evolución de la vida. Los eucariotas y, en particular, los akaryotes (Bacteria y Archaea) evolucionaron a través de la pérdida reductiva, de modo que las similitudes resultan de la retención diferencial de las características originales.
Suponiendo que no haya otro grupo involucrado, hay tres filogenias posibles para las bacterias, arqueas y eucariotas en las que cada una es monofilética. Estos están etiquetados de 1 a 3 en la tabla a continuación. La hipótesis de los eocytes es una modificación de la hipótesis 2 en la cual las Archaea son parafiléticas. (La tabla y los nombres de las hipótesis se basan en Harish y Kurland, 2017.)
| 1 - Dos imperios | 2 - Tres dominios | 3 - Gupta | 4 - Eocito | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
|
|
|
En los últimos años, la mayoría de los investigadores han favorecido los tres dominios (3D) o las hipótesis de los eocytes. Un análisis de ARNr apoya el escenario de eocitos, aparentemente con la raíz eucariota en Excavata. Un cladograma que respalda la hipótesis de los eocytes, colocando eucariotas dentro de Archaea, basado en análisis filogenómicos de Asgard archaea, es:
| Proteoarchaeota |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||
En este escenario, el grupo Asgard es visto como un taxón hermano del grupo TACK, que comprende Crenarchaeota (antes llamado eocitos), Thaumarchaeota y otros.
En 2017, hubo un retroceso significativo contra este escenario, argumentando que los eucariotas no surgieron dentro de las Archaea. Cunha et al. análisis producidos que apoyan la hipótesis de los tres dominios (3D) o de Woese (2 en la tabla anterior) y rechazan la hipótesis de los evocitos (4 arriba). Harish y Kurland encontraron un fuerte apoyo para los dos imperios anteriores (2D) o la hipótesis de Mayr (1 en la tabla anterior), en base al análisis de las secuencias codificantes de los dominios proteicos. Rechazaron la hipótesis de los evocitos como la menos probable. Una posible interpretación de su análisis es que el ancestro común universal (UCA) del árbol de la vida actual era un organismo complejo que sobrevivió a un cuello de botella evolutivo, en lugar de un organismo más simple que surgió temprano en la historia de la vida.
Sistema de endomembrana y mitocondrias
Los orígenes del sistema de endomembrana y las mitocondrias tampoco están claros. La hipótesis phagotrófica propone que las membranas de tipo eucariótico que carecen de una pared celular se originaron primero, con el desarrollo de endocitosis, mientras que las mitocondrias se adquirieron por ingestión como endosimbiontes. La hipótesis sintrófica propone que el proto-eucariota se basó en la proto-mitocondria para obtener alimento, y así creció para rodearlo. Aquí las membranas se originaron después del englobamiento de la mitocondria, en parte gracias a los genes mitocondriales (la hipótesis del hidrógeno es una versión particular).
En un estudio que usa genomas para construir supertrees, Pisani et al. (2007) sugieren que, junto con la evidencia de que nunca hubo un eucariota sin mitocondria, los eucariotas evolucionaron a partir de una sintrofia entre una archaea estrechamente relacionada con Thermoplasmatales y una α-proteobacteria, probablemente una simbiosis impulsada por azufre o hidrógeno. La mitocondria y su genoma es un remanente del endosimbionte α-proteobacterial.
Hipótesis
Se han propuesto diferentes hipótesis sobre cómo nacieron las células eucarióticas. Estas hipótesis se pueden clasificar en dos clases distintas: modelos autógenos y modelos quiméricos.
Modelos autógenos
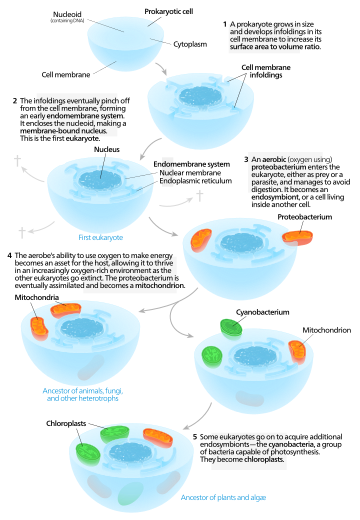
Los modelos autógenos proponen que una célula proto-eucariota que contiene un núcleo existió primero y más tarde adquirió mitocondrias. Según este modelo, un procariota grande desarrolló invaginaciones en su membrana plasmática con el fin de obtener suficiente área superficial para mantener su volumen citoplásmico. A medida que las invaginaciones se diferenciaron en función, algunas se convirtieron en compartimentos separados, dando lugar al sistema de endomembrana, que incluye el retículo endoplasmático, el aparato de Golgi, la membrana nuclear y las estructuras de una sola membrana, como los lisosomas. Se propone que las mitocondrias provienen de la endosimbiosis de una proteobacteria aeróbica, y se supone que todos los linajes eucariotas que no adquirieron mitocondrias se extinguieron. Los cloroplastos surgieron de otro evento endosimbiótico que involucraba cianobacterias. Dado que todos los eucariontes tienen mitocondrias,
Modelos quiméricos
Los modelos quiméricos afirman que inicialmente existían dos células procariotas: una archaeon y una bacteria. Estas células se sometieron a un proceso de fusión, ya sea por fusión física o por endosimbiosis, lo que conduce a la formación de una célula eucariota. Dentro de estos modelos quiméricos, algunos estudios afirman además que las mitocondrias se originaron a partir de un ancestro bacteriano, mientras que otros enfatizan el papel de los procesos endosimbióticos detrás del origen de las mitocondrias.
Con base en el proceso de simbiosis mutualista, las hipótesis se pueden categorizar como - la teoría endosimbiótica en serie (SET), la hipótesis del hidrógeno (principalmente un proceso de simbiosis donde la transferencia de hidrógeno tiene lugar entre diferentes especies) y la hipótesis de la sintrofia.
De acuerdo con la teoría endosimbiótica en serie (defendida por Lynn Margulis), una unión entre una bacteria anaeróbica móvil (como Spirochaeta ) y una crenarchaeon termoacidophilic (como Thermoplasma que es de naturaleza sulfidogénica) dio lugar a los eucariotas actuales. Esta unión estableció un organismo móvil capaz de vivir en las aguas ácidas y sulfurosas ya existentes. Se sabe que el oxígeno causa toxicidad a organismos que carecen de la maquinaria metabólica requerida. Por lo tanto, la archaeon proporcionó a la bacteria un entorno reducido altamente beneficioso (el azufre y el sulfato se redujeron a sulfuro). En condiciones microaerófilas, el oxígeno se redujo a agua, creando así una plataforma de beneficio mutuo. La bacteria, por otro lado, aportó los productos de fermentación necesarios y los aceptores de electrones junto con su característica de movilidad a la archaeon, obteniendo así una motilidad de natación para el organismo. De un consorcio de ADN bacteriano y arcaico se originó el genoma nuclear de células eucariotas. Spirochetesgave aumentan las características móviles de las células eucarióticas. Las unificaciones endosimbióticas de los ancestros de las alfa-proteobacterias y las cianobacterias, condujeron al origen de las mitocondrias y los plástidos, respectivamente. Por ejemplo, Se sabe que el tiodendronse originó a través de un proceso ectosimbiótico basado en una sintrofia similar de azufre existente entre los dos tipos de bacterias: Desulphobacter y Spirochaeta . Sin embargo, tal asociación basada en la simbiosis móvil nunca se ha observado prácticamente. Además, no hay evidencia de arqueas y espiroquetas que se adapten a ambientes intensos basados en ácido.
En la hipótesis del hidrógeno, el enlace simbiótico de una archaeon metanógena (hospedador) anaeróbica y autótrofa con una alfa-proteobacteria (el simbionte) dio lugar a los eucariotas. El hidrógeno huésped utilizado (H 2 ) y dióxido de carbono (CO 2 ) para producir metano mientras que el simbionte, capaz de la respiración aeróbica, expulsado H 2 y CO 2 como subproductos del proceso de fermentación anaerobia. El ambiente metanogénico del anfitrión funcionó como un sumidero de H 2, lo que resultó en una mayor fermentación bacteriana. La transferencia génica endosimbiótica (EGT) actuó como un catalizador para que el huésped adquiriera el metabolismo de carbohidratos de los simbiontes y se volviera heterotrófico en la naturaleza. Posteriormente, se perdió la capacidad de formación de metano del huésped. Por lo tanto, los orígenes del orgánulo heterótrofo (simbionte) son idénticos a los orígenes del linaje eucariota. En esta hipótesis, la presencia de H 2 representa la fuerza selectiva que forjó eucariotas de procariotas.
La hipótesis de la sintrofia se desarrolló en contraste con la hipótesis del hidrógeno y propone la existencia de dos eventos simbióticos. De acuerdo con esta teoría, el origen de las células eucarióticas se basó en la simbiosis metabólica (sintrofia) entre una archaeon metanogénica y una delta-proteobacterium. Esta simbiosis sintrófica fue inicialmente facilitada por H 2 transferencia entre diferentes especies en ambientes anaeróbicos. En etapas anteriores, una alfa-proteobacteria se convirtió en un miembro de esta integración, y más tarde se convirtió en la mitocondria. La transferencia de genes de una delta-proteobacterium a una archaeon condujo a la archaeon metanogénica que se convertía en un núcleo. La arquea constituyó el aparato genético, mientras que la delta-proteobacteria contribuyó a las características citoplásmicas. Esta teoría incorpora dos fuerzas selectivas en el momento de la evolución del núcleo: (a) presencia de partición metabólica para evitar los efectos nocivos de la coexistencia de vías celulares anabólicas y catabólicas, y (b) prevención de la biosíntesis anormal de proteínas debido a una gran propagación de intrones en los genes archaeal después de adquirir la mitocondria y perder la metanogénesis.
Obtenido de: https://en.wikipedia.org/wiki/Eukaryote