Archaea
Definición
| Archaea (Archaebacteria) Rango temporal: 3.5-0Ga Had'n Archean Proterozoico Pha. Paleoarqueano o tal vez Eoarchean - reciente | |
|---|---|
 | |
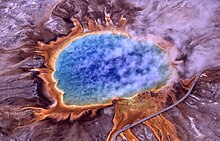
| Halobacterium sp. cepa NRC-1, cada celda de aproximadamente 5 μm de largo | |
| clasificación cientifica | |
| Dominio: | Archaea Woese, Kandler y Wheelis, 1990 |
| Reinos y phyla | |
| |
| Sinónimos | |
| |
Las células de Archaeal tienen propiedades únicas que las separan de los otros dos dominios de la vida, Bacteria y Eukarya. Las Archaea se dividen en múltiples phyla reconocidos. La clasificación es difícil porque la mayoría no se ha aislado en el laboratorio y solo se ha detectado mediante el análisis de sus ácidos nucleicos en muestras de su entorno.
Las arqueas y las bacterias son generalmente similares en tamaño y forma, aunque algunas arqueas tienen formas bastante diferentes a las de las bacterias, como las células planas y cuadradas de Haloquadratum walsbyi.. A pesar de esta similitud morfológica con las bacterias, las arqueas poseen genes y varias rutas metabólicas que están más estrechamente relacionadas con las de los eucariotas, especialmente las enzimas involucradas en la transcripción y la traducción. Otros aspectos de la bioquímica arquearia son únicos, como su dependencia de los lípidos de éter en sus membranas celulares, incluidos los arqueoles. Las arqueas usan más fuentes de energía que las eucariotas: varían desde compuestos orgánicos, como azúcares, hasta amoníaco, iones metálicos o incluso gas de hidrógeno. Las arqueas tolerantes a la sal (Haloarchaea) usan la luz del sol como fuente de energía, y otras especies de archaea fijan carbono, pero a diferencia de las plantas y las cianobacterias, ninguna especie conocida de arqueas hace ambas cosas. Las arqueas se reproducen asexualmente por fisión binaria, fragmentación o gemación; a diferencia de las bacterias y eucariotas, ninguna especie conocida forma esporas.
Las arqueas fueron inicialmente vistas como extremófilos que viven en ambientes hostiles, como aguas termales y lagos salados, pero desde entonces se han encontrado en una amplia gama de hábitats, incluidos suelos, océanos y marismas. También son parte de la microbiota humana, que se encuentra en el colon, la boca y la piel. Las Arqueas son particularmente numerosas en los océanos, y las arqueas en el plancton pueden ser uno de los grupos más abundantes de organismos en el planeta. Las arqueas son una parte importante de la vida de la Tierra y pueden desempeñar funciones tanto en el ciclo del carbono como en el del nitrógeno. No se conocen ejemplos claros de patógenos arcaicos o parásitos, pero a menudo son mutualistas o comensales. Un ejemplo son los metanógenos que habitan las tripas de humanos y rumiantes, donde su gran cantidad ayuda a la digestión. Los metanógenos también se usan en la producción de biogás y en el tratamiento de aguas residuales.
Clasificación
Nuevo dominio
Durante gran parte del siglo XX, los procariotas fueron considerados como un solo grupo de organismos y clasificados en función de su bioquímica, morfología y metabolismo. Por ejemplo, los microbiólogos trataron de clasificar los microorganismos basándose en las estructuras de sus paredes celulares, sus formas y las sustancias que consumen. En 1965, Emile Zuckerkandl y Linus Pauling propusieron utilizar las secuencias de los genes en diferentes procariotas para determinar cómo están relacionados entre sí. Este enfoque filogenético es el principal método utilizado en la actualidad.
Las arqueas se clasificaron por primera vez como un grupo separado de procariotas en 1977 por Carl Woese y George E. Fox basándose en las secuencias de los genes del ARN ribosomal (ARNr). Estos dos grupos fueron originalmente llamados Archaebacteria y Eubacteria y se denominaron Urkingdoms por Woese y Fox; otros investigadores los trataron como reinos o subjuegos. En esta publicación, Woese y Fox presentaron las primeras pruebas para el concepto de arqueobacteria, que en ese momento contenía solo los metanógenos, como una "línea de descenso" separada: 1. falta de peptidoglicano en sus paredes celulares, 2. dos coenzimas inusuales, 3. resultados de la secuenciación del gen del ARN ribosómico 16S. Para enfatizar esta diferencia, Woese, Otto Kandler y Mark Wheelis más tarde propusieron un nuevo sistema natural de organismos con tres dominios separados: el Eukarya, las Bacterias y las Archaea, en lo que ahora se conoce como "La Revolución Woesiana". La historia de la investigación sobre Archaea está documentada en detalle por Jan Sapp.
La palabra archaea proviene del griego clásico ἀρχαῖα, que significa "cosas antiguas", ya que los primeros representantes del dominio Archaea eran metanógenos y se suponía que su metabolismo reflejaba la atmósfera primitiva de la Tierra y la antigüedad de los organismos, pero a medida que se estudiaban nuevos hábitats, se descubrieron más organismos. Los microbios extremos halófilos e hipertermófilos también se incluyeron en Archaea. Durante mucho tiempo, las arqueas fueron vistas como extremófilos que solo existen en hábitats extremos tales como aguas termales y lagos salados. A fines del siglo XX, las arqueas también habían sido identificadas en ambientes no extremos. Hoy en día, se sabe que son un grupo grande y diverso de organismos que están ampliamente distribuidos en la naturaleza y son comunes en todos los hábitats. Esta nueva apreciación de la importancia y ubicuidad de las arqueas proviene del uso de la reacción en cadena de la polimerasa (PCR) para detectar procariotas de muestras ambientales (como agua o suelo) al multiplicar sus genes ribosómicos. Esto permite la detección e identificación de organismos que no han sido cultivados en el laboratorio.
Clasificación actual

La clasificación de las arqueas y de los procariotas en general es un campo polémico y en constante movimiento. Los sistemas actuales de clasificación pretenden organizar arqueas en grupos de organismos que comparten características estructurales y ancestros comunes. Estas clasificaciones dependen en gran medida del uso de la secuencia de genes de ARN ribosómico para revelar las relaciones entre organismos (filogenia molecular). La mayoría de las especies cultivables y bien investigadas de arqueas son miembros de dos phyla principales, Euryarchaeota y Crenarchaeota. Otros grupos han sido tentativamente creados. Por ejemplo, la peculiar especie Nanoarchaeum equitans, que fue descubierto en 2003, recibió su propio phylum, el Nanoarchaeota. También se ha propuesto un nuevo phylum Korarchaeota. Contiene un pequeño grupo de especies termófilas inusuales que comparte características de los dos phyla principales, pero está más estrechamente relacionado con la Crenarchaeota. Otras especies de arqueas detectadas recientemente están relacionadas lejanamente con cualquiera de estos grupos, como los nanoorganismos acidófilos de la Archaeal Richmond Mine (ARMAN, que comprenden Micrarchaeota y Parvarchaeota), que se descubrieron en 2006 y son algunos de los organismos más pequeños conocidos.
Se ha propuesto un superphylum - TACK - que incluye el Thaumarchaeota, Aigarchaeota, Crenarchaeota y Korarchaeota. Este superphylum puede estar relacionado con el origen de eucariotas. Más recientemente, el superfilo Asgard ha sido nombrado y propuesto para estar más estrechamente relacionado con el eucariota original y un grupo hermano de TACK.Concepto de especie
La clasificación de arqueas en especies también es controvertida. La biología define una especie como un grupo de organismos relacionados. El criterio de crianza exclusivo familiar (organismos que pueden reproducirse entre sí pero no con otros) no sirve de nada porque las arqueas se reproducen asexualmente.
Las arqueas muestran altos niveles de transferencia génica horizontal entre linajes. Algunos investigadores sugieren que los individuos se pueden agrupar en poblaciones similares a especies que poseen genomas muy similares y una transferencia de genes infrecuente a / de células con genomas menos relacionados, como en el género Ferroplasma . Por otro lado, los estudios en Halorubrum encontraron una transferencia genética significativa hacia / desde poblaciones menos relacionadas, lo que limita la aplicabilidad del criterio. Una segunda preocupación es hasta qué punto tales designaciones de especies tienen un significado práctico.
El conocimiento actual sobre la diversidad genética es fragmentario y el número total de especies de archaeas no se puede estimar con precisión. Las estimaciones del número de phyla oscilan entre 18 y 23, de las cuales solo 8 tienen representantes que se han cultivado y estudiado directamente. Muchos de estos grupos hipotéticos se conocen a partir de una única secuencia de ARNr, lo que indica que la diversidad entre estos organismos sigue siendo desconocida. Las bacterias también contienen muchos microbios incultos con implicaciones similares para la caracterización.
En promedio, las secuencias de ADN de archaeal (genoma completo) muestran niveles de complejidad más altos que los de las bacterias.
Origen y evolución
La edad de la Tierra es de unos 4.540 millones de años. La evidencia científica sugiere que la vida comenzó en la Tierra hace al menos 3.5 mil millones de años. La primera evidencia de vida en la Tierra es el grafito que se descubrió biogénico en rocas metasedimentarias de 3.700 millones de años descubiertas en el oeste de Groenlandia y fósiles de esteras microbianas halladas en areniscas de 3.48 billones de años descubiertas en Australia Occidental. Más recientemente, en 2015, se encontraron "restos de vida biótica" en rocas de 4,100 millones de años en el oeste de Australia.
Aunque los probables fósiles de células procariotas datan de hace casi 3.500 millones de años, la mayoría de los procariotas no tienen morfologías distintivas y las formas fósiles no pueden utilizarse para identificarlos como arqueas. En cambio, los fósiles químicos de lípidos únicos son más informativos porque tales compuestos no ocurren en otros organismos. Algunas publicaciones sugieren que restos de lípidos arcaicos o eucariotas están presentes en las lutitas que datan de hace 2.700 millones de años; tales datos han sido cuestionados desde entonces. Dichos lípidos también se han detectado en rocas aún más antiguas del oeste de Groenlandia. Las huellas más antiguas provienen del distrito de Isua, que incluye los sedimentos conocidos más antiguos de la Tierra, formados hace 3.800 millones de años. El linaje archaeal puede ser el más antiguo que existe en la Tierra.
Woese argumentó que las bacterias, las arqueas y las eucariotas representan líneas de descendencia separadas que divergían desde el principio de una colonia ancestral de organismos. Una posibilidad es que esto ocurrió antes de la evolución de las células, cuando la falta de una membrana celular típica permitió la transferencia génica lateral no restringida, y que los ancestros comunes de los tres dominios surgieron por la fijación de subconjuntos específicos de genes. Es posible que el último ancestro común de las bacterias y arqueas era un termófilo, lo que plantea la posibilidad de que las temperaturas más bajas son las "condiciones extremas" en términos de arqueas y organismos que viven en ambientes más frescos sólo aparecieron más tarde. Como las arqueas y las bacterias no están más relacionadas entre sí que entre eucariotas, el término procariotaEl único significado que sobrevive es "no es un eucariota", lo que limita su valor.
Comparación con otros dominios
La siguiente tabla compara algunas características principales de los tres dominios, para ilustrar sus similitudes y diferencias. Muchas de estas características también se discuten a continuación.
| Propiedad | Archaea | Bacteria | Eukarya |
|---|---|---|---|
| Membrana celular | Lípidos ligados a éter, pseudopeptidoglucano | Lípidos ligados a éster, peptidoglucano | Lípidos ligados a éster, diversas estructuras |
| Estructura génica | Cromosomas circulares, traducción y transcripción similares a Eukarya | Cromosomas circulares, traducción única y transcripción | Múltiples cromosomas lineales, traducción y transcripción similares a Archaea |
| Estructura interna | No hay orgánulos unidos a la membrana (cuestionados) o núcleo | No hay organelos o núcleos unidos a la membrana | Organelos y núcleos unidos a la membrana |
| Metabolismo | Varios, con metanogénesis exclusiva de Archaea | Varios, incluida la fotosíntesis, la respiración aeróbica y anaeróbica, la fermentación y la autotrofía | Fotosíntesis, respiración celular y fermentación |
| Reproducción | Reproducción asexual, transferencia horizontal de genes | Reproducción asexual, transferencia horizontal de genes | Reproducción sexual y asexual |
Archaea se separó como un tercer dominio debido a las grandes diferencias en su estructura de ARN ribosomal. La molécula particular de ARN secuenciada, conocida como ARNr 16S, está presente en todos los organismos y siempre tiene la misma función vital: la producción de proteínas. Debido a que esta función es tan importante para la vida, es poco probable que los organismos con mutaciones de su ARNr 16S sobrevivan, lo que conduce a una gran estabilidad en la estructura de este nucleótido a lo largo de muchas generaciones. El ARNr 16S también es lo suficientemente grande como para retener información específica del organismo, pero lo suficientemente pequeña para ser secuenciada en un tiempo manejable. En 1977, Carl Woese, un microbiólogo que estudiaba la secuenciación genética de organismos, desarrolló un nuevo método de secuenciación que implicaba dividir el ARN en fragmentos que podían clasificarse y compararse con otros fragmentos de otros organismos.
Woese usó su nuevo método de comparación de ARNr para categorizar y contrastar diferentes organismos. Secuenció una variedad de diferentes especies y se topó con un grupo de metanógenos que tenían patrones muy diferentes a los conocidos como procariotas o eucariotas. Estos metanógenos fueron mucho más similares entre sí que lo que fueron para otros organismos secuenciados, lo que llevó a Woese a proponer el nuevo dominio de Archaea. Sus experimentos demostraron que las Archaea eran más similares a las eucariotas que las procariotas, a pesar de que eran más similares a las estructuras procariotas. Esto llevó a la conclusión de que Archaea y Eukarya compartían un ancestro común más reciente que Eukarya y Bacteria en general. El desarrollo del núcleo se produjo después de la división entre las bacterias y este ancestro común. Aunque las Archaea son procariotas,
Una propiedad exclusiva de Archaea es el uso abundante de lípidos ligados a éter en sus membranas celulares. Los enlaces de éter son químicamente más estables que los enlaces éster encontrados en Bacteria y Eukarya, que pueden ser un factor que contribuye a la capacidad de muchas Archaea para sobrevivir en ambientes extremos que ejercen una gran presión sobre las membranas celulares, como el calor extremo y la salinidad. El análisis comparativo de los genomas de arqueas también ha identificado varias firmas moleculares en forma de indeles de firma conservados y proteínas de firma que están presentes de manera única en todas las Archaea o en los diferentes grupos principales dentro de Archaea. Otra característica única de Archaea es que ningún otro organismo conocido es capaz de metanogénesis (la producción metabólica de metano). Las arqueas metanogénicas desempeñan un papel fundamental en los ecosistemas con organismos que obtienen energía de la oxidación del metano, muchas de las cuales son bacterias, ya que a menudo son una fuente importante de metano en dichos entornos y pueden desempeñar un papel como productores primarios. Los metanógenos también juegan un papel crítico en el ciclo del carbono, descomponiendo el carbono orgánico en metano, que también es un importante gas de efecto invernadero.
Relación con las bacterias

La relación entre los tres dominios es de importancia central para comprender el origen de la vida. La mayoría de las vías metabólicas, que son el objeto de la mayoría de los genes de un organismo, son comunes entre Archaea y Bacteria, mientras que la mayoría de los genes implicados en la expresión del genoma son comunes entre Archaea y Eukarya. Dentro de los procariotas, la estructura de las células arqueales es más similar a la de las bacterias gram-positivas, en gran parte porque ambas tienen una sola bicapa lipídica y generalmente contienen un sacáculo grueso (exoesqueleto) de composición química variable. En algunos árboles filogenéticos basados en diferentes secuencias de genes / proteínas de homólogos procariotas, los homólogos de arqueas están más estrechamente relacionados con los de bacterias gram-positivas. Archaea y bacterias grampositivas también comparten indeles conservados en varias proteínas importantes,
Se ha propuesto que las arqueas evolucionaron a partir de bacterias grampositivas en respuesta a la presión de selección de antibióticos. Esto es sugerido por la observación de que las arqueas son resistentes a una amplia variedad de antibióticos que se producen principalmente por bacterias gram-positivas, y que estos antibióticos actúan principalmente sobre los genes que distinguen a las arqueas de las bacterias. La propuesta es que la presión selectiva hacia la resistencia generada por los antibióticos gram-positivos finalmente fue suficiente para causar cambios extensos en muchos de los genes diana de los antibióticos, y que estas cepas representaron los ancestros comunes de las actuales arqueas. La evolución de Archaea en respuesta a la selección de antibióticos, o cualquier otra presión competitiva selectiva, también podría explicar su adaptación a ambientes extremos (como la alta temperatura o la acidez) como resultado de la búsqueda de nichos desocupados para escapar de organismos productores de antibióticos; Cavalier-Smith ha hecho una sugerencia similar. Esta propuesta también es apoyada por otro trabajo que investiga las relaciones estructurales proteínicas y estudios que sugieren que las bacterias grampositivas pueden constituir los primeros linajes de ramificación dentro de los procariotas.
Relación con eucariotas
La relación evolutiva entre arqueas y eucariotas sigue sin estar clara. Además de las similitudes en la estructura y función celular que se discuten a continuación, muchos árboles genéticos agrupan a los dos.
Los factores complicados incluyen afirmaciones de que la relación entre eucariotas y el arilo filo Crenarchaeota es más estrecha que la relación entre Euryarchaeota y el phylum Crenarchaeota y la presencia de genes similares a arqueas en ciertas bacterias, como Thermotoga maritima , de la transferencia horizontal de genes. La hipótesis estándar establece que el ancestro de los eucariotas divergió temprano de las Archaea, y que los eucariotas surgieron a través de la fusión de una arquea y eubacteria, que se convirtieron en el núcleo y el citoplasma; esto explica varias similitudes genéticas, pero tiene dificultades para explicar la estructura celular. Una hipótesis alternativa, la hipótesis de los eocitos, postula que Eukaryota surgió relativamente tarde de las Archaea.
Un linaje de arqueas descubierto en 2015, Lokiarchaeum (del nuevo Phylum propuesto "Lokiarchaeota"), llamado así por un respiradero hidrotermal llamado Castillo de Loki en el Océano Ártico, se ha encontrado que está más estrechamente relacionado con los eucariotas conocidos en este momento. Se lo ha llamado un organismo de transición entre procariotas y eucariotas.
Se han encontrado varios phyla hermanos de "Lokiarchaeota" ("Thorarchaeota", "Odinarchaeota", "Heimdallarchaeota"), todos juntos que comprenden un Asgard supergrupo recién propuesto, que puede aparecer como un taxón hermano de TACK. Los detalles de la relación de los miembros de Asgard y los eucariotas aún se están considerando.
Morfología
Las arqueas individuales varían desde 0.1 micrómetros (μm) hasta más de 15 μm de diámetro, y ocurren en varias formas, comúnmente como esferas, varillas, espirales o placas. Otras morfologías en la Crenarchaeota incluyen células lobuladas de forma irregular en Sulfolobus , filamentos en forma de aguja que tienen menos de medio micrómetro de diámetro en Thermofilum , y varillas casi perfectamente rectangulares en Thermoproteus y Pyrobaculum . Archaea en el género Haloquadratum como Haloquadratum walsbyi son arqueas planas y cuadradas que viven en piscinas hipersalinas. Estas formas inusuales son probablemente mantenidas tanto por sus paredes celulares como por un citoesqueleto procariótico. Las proteínas relacionadas con los componentes del citoesqueleto de otros organismos existen en arqueas, y los filamentos se forman dentro de sus células, pero a diferencia de otros organismos, estas estructuras celulares son poco conocidas. En Thermoplasma y Ferroplasma, la falta de una pared celular significa que las células tienen formas irregulares y pueden parecerse a las amebas.
Algunas especies forman agregados o filamentos de células de hasta 200 μm de largo. Estos organismos pueden ser prominentes en las biopelículas. Notablemente, los agregados de células de Thermococcus coalescens se fusionan en cultivo, formando células gigantes únicas. Archaea en el género Pyrodictium produce una colonia multicell elaborada que involucra matrices de tubos huecos largos y delgados llamados cánulas que sobresalen de las superficies de las células y las conectan a una aglomeración densa similar a un arbusto. La función de estas cánulas no está establecida, pero pueden permitir la comunicación o el intercambio de nutrientes con los vecinos. Existen colonias multiespecíficas, como la comunidad de "cadena de perlas" que se descubrió en 2001 en un pantano alemán. Las colonias redondas y blanquecinas de una nueva especie de Euryarchaeota están espaciadas a lo largo de finos filamentos que pueden llegar a medir hasta 15 centímetros (5,9 pulgadas) de largo; estos filamentos están hechos de una especie de bacteria particular.
Estructura, desarrollo de composición y operación
Las arqueas y las bacterias generalmente tienen una estructura celular similar, pero la composición y la organización de las células hacen que las arqueas se separen. Al igual que las bacterias, las arqueas carecen de membranas interiores y orgánulos. Al igual que las bacterias, las membranas celulares de las arqueas generalmente están limitadas por una pared celular y nadan usando uno o más flagelos. Estructuralmente, las arqueas son más similares a las bacterias gram-positivas. La mayoría tienen una sola membrana plasmática y pared celular, y carecen de espacio periplásmico; la excepción a esta regla general es Ignicoccus , que posee un periplasma particularmente grande que contiene vesículas unidas a la membrana y está encerrado por una membrana externa.
Pared celular y flagelos
La mayoría de las arqueas (pero no Thermoplasma y Ferroplasma ) poseen una pared celular. En la mayoría de las arqueas, la pared se ensambla a partir de proteínas de la capa superficial, que forman una capa S. Una capa S es una matriz rígida de moléculas de proteínas que cubren el exterior de la célula (como el correo en cadena). Esta capa proporciona protección química y física, y puede evitar que las macromoléculas entren en contacto con la membrana celular. A diferencia de las bacterias, las arqueas carecen de peptidoglicano en sus paredes celulares. Las metanobacteriales tienen paredes celulares que contienen pseudopeptidoglicano, que se asemeja al peptidoglicano eubacterial en morfología, función y estructura física, pero el pseudopeptidoglicano es distinto en la estructura química; carece de D-aminoácidos y ácido N-acetilmurámico.
Los archaea flagella funcionan como flagelos bacterianos, sus largos tallos son impulsados por motores rotatorios en la base. Estos motores son alimentados por el gradiente de protones a través de la membrana, pero los flagelos archaeal son notablemente diferentes en composición y desarrollo. Los dos tipos de flagelos evolucionaron a partir de diferentes ancestros. El flagelo bacteriano comparte un ancestro común con el sistema de secreción de tipo III, mientras que los flagelos arquetípicos parecen haber evolucionado a partir de pili de tipo IV bacteriano. En contraste con el flagelo bacteriano, que es hueco y está ensamblado por subunidades que se mueven hacia arriba por el poro central hasta la punta de los flagelos, los flagelos arqueados se sintetizan mediante la adición de subunidades en la base.
Membranas

Las membranas Arqueales están hechas de moléculas que son claramente diferentes de las de todas las demás formas de vida, lo que demuestra que las arqueas se relacionan solo remotamente con bacterias y eucariotas. En todos los organismos, las membranas celulares están hechas de moléculas conocidas como fosfolípidos. Estas moléculas poseen tanto una parte polar que se disuelve en el agua (la "cabeza" del fosfato), como una parte no polar "grasosa" que no (la cola de los lípidos). Estas partes diferentes están conectadas por un resto de glicerol. En el agua, los fosfolípidos se agrupan, con las cabezas mirando hacia el agua y las colas mirando hacia afuera. La estructura principal en las membranas celulares es una doble capa de estos fosfolípidos, que se llama una bicapa lipídica.
Los fosfolípidos de las arqueas son inusuales en cuatro formas:
- Tienen membranas compuestas de lípidos de glicerol-éter, mientras que las bacterias y eucariotas tienen membranas compuestas principalmente de lípidos de ésteres de glicerol. La diferencia es el tipo de enlace que une los lípidos al resto de glicerol; los dos tipos se muestran en amarillo en la figura de la derecha. En los lípidos éster, este es un enlace éster, mientras que en los lípidos éter, este es un enlace éter. Los enlaces éter son químicamente más resistentes que los enlaces éster.
- La estereoquímica del resto de glicerol arqueado es la imagen especular de la que se encuentra en otros organismos. El resto glicerol puede aparecer en dos formas que son imágenes especulares entre sí, llamadas enantiómeros . Del mismo modo que una mano derecha no encaja fácilmente en un guante zurdo, los enantiómeros de un tipo generalmente no pueden usarse ni fabricarse mediante enzimas adaptadas para el otro. Los fosfolípidos archaeal se basan en una columna vertebral de sn -glicerol-1-fosfato, que es un enantiómero de sn-glicerol-3-fosfato, el esqueleto de fosfolípidos encontrado en bacterias y eucariotas. Esto sugiere que las arqueas usan enzimas completamente diferentes para sintetizar fosfolípidos que las bacterias y eucariotas. Tales enzimas se desarrollaron muy temprano en la historia de la vida, lo que indica una división temprana de los otros dos dominios.
- Las colas de lípidos de Archaeal difieren de las de otros organismos en que se basan en largas cadenas isoprenoides con múltiples ramas laterales, a veces con anillos de ciclopropano o ciclohexano. Por el contrario, los ácidos grasos en las membranas de otros organismos tienen cadenas rectas sin ramas laterales o anillos. Aunque los isoprenoides desempeñan un papel importante en la bioquímica de muchos organismos, solo las arqueas los usan para producir fosfolípidos. Estas cadenas ramificadas pueden ayudar a evitar que las membranas archaeas se filtren a altas temperaturas.
- En algunas arqueas, la bicapa lipídica se reemplaza por una monocapa. En efecto, las arqueas fusionan las colas de dos moléculas de fosfolípidos en una sola molécula con dos cabezas polares (un bolaamphiphile); esta fusión puede hacer que sus membranas sean más rígidas y más capaces de resistir ambientes hostiles. Por ejemplo, los lípidos en Ferroplasma son de este tipo, que se cree que ayudan a la supervivencia de este organismo en su hábitat altamente ácido.
Metabolismo
Las arqueas exhiben una gran variedad de reacciones químicas en su metabolismo y usan muchas fuentes de energía. Estas reacciones se clasifican en grupos nutricionales, según la energía y las fuentes de carbono. Algunas arqueas obtienen energía de compuestos inorgánicos como azufre o amoníaco (son litotrofos). Estos incluyen nitrificadores, metanógenos y metanoxicadores anaeróbicos. En estas reacciones, un compuesto pasa electrones a otro (en una reacción redox), liberando energía para alimentar las actividades de la célula. Un compuesto actúa como un donante de electrones y uno como un aceptor de electrones. La energía liberada se utiliza para generar trifosfato de adenosina (ATP) a través de la quimiosmosis, el mismo proceso básico que ocurre en la mitocondria de las células eucarióticas.
Otros grupos de arqueas usan la luz del sol como fuente de energía (son fotótrofos), pero la fotosíntesis generadora de oxígeno no ocurre en ninguno de estos organismos. Muchas vías metabólicas básicas se comparten entre todas las formas de vida; por ejemplo, las arqueas usan una forma modificada de glucólisis (la vía Entner-Doudoroff) y un ciclo completo o parcial de ácido cítrico. Estas similitudes con otros organismos probablemente reflejan los orígenes tempranos en la historia de la vida y su alto nivel de eficiencia.
| Tipo nutricional | Fuente de energía | Fuente de carbono | Ejemplos |
|---|---|---|---|
| Fototrofos | Luz de sol | Compuestos orgánicos | Halobacterium |
| Lithotrophs | Compuestos inorgánicos | Compuestos orgánicos o fijación de carbono | Ferroglobus , Methanobacteria o Pyrolobus |
| Organotrofos | Compuestos orgánicos | Compuestos orgánicos o fijación de carbono | Pyrococcus , Sulfolobus o Methanosarcinales |
Algunos Euryarchaeota son metanógenos (arqueas que producen metano como resultado del metabolismo) que viven en ambientes anaeróbicos, como los pantanos. Esta forma de metabolismo evolucionó temprano, e incluso es posible que el primer organismo de vida libre fuera un metanogeno. Una reacción común implica el uso de dióxido de carbono como un aceptor de electrones para oxidar el hidrógeno. La metanogénesis involucra un rango de coenzimas que son únicas a estas arqueas, como la coenzima M y el metanofurano. Otros compuestos orgánicos tales como alcoholes, ácido acético o ácido fórmico se usan como aceptores de electrones alternativos por metanógenos. Estas reacciones son comunes en las arqueas que habitan el intestino. El ácido acético también se descompone en metano y dióxido de carbono directamente, por acetotrophic Archaea. Estos acetotropos son arqueas en el orden Methanosarcinales, y son una parte importante de las comunidades de microorganismos que producen biogás.

Otras arqueas usan CO
2 en la atmósfera como fuente de carbono, en un proceso llamado fijación de carbono (son autótrofos). Este proceso implica una forma altamente modificada del ciclo de Calvin o una vía metabólica descubierta recientemente llamada ciclo 3-hidroxipropionato / 4-hidroxibutirato. Los Crenarchaeota también usan el ciclo inverso de Krebs mientras que los Euryarchaeota también usan la vía reductiva de acetil-CoA. La fijación de carbono es alimentada por fuentes de energía inorgánica. No hay arqueas conocidas que realicen la fotosíntesis. Las fuentes de energía Arqueal son extremadamente diversas, y van desde la oxidación del amoníaco por el Nitrosopumilales hasta la oxidación del sulfuro de hidrógeno o azufre elemental por especies de Sulfolobus, utilizando oxígeno o iones metálicos como aceptores de electrones.
2 en la atmósfera como fuente de carbono, en un proceso llamado fijación de carbono (son autótrofos). Este proceso implica una forma altamente modificada del ciclo de Calvin o una vía metabólica descubierta recientemente llamada ciclo 3-hidroxipropionato / 4-hidroxibutirato. Los Crenarchaeota también usan el ciclo inverso de Krebs mientras que los Euryarchaeota también usan la vía reductiva de acetil-CoA. La fijación de carbono es alimentada por fuentes de energía inorgánica. No hay arqueas conocidas que realicen la fotosíntesis. Las fuentes de energía Arqueal son extremadamente diversas, y van desde la oxidación del amoníaco por el Nitrosopumilales hasta la oxidación del sulfuro de hidrógeno o azufre elemental por especies de Sulfolobus, utilizando oxígeno o iones metálicos como aceptores de electrones.
Las arqueas fototróficas usan la luz para producir energía química en forma de ATP. En las Halobacterias, las bombas de iones activadas por la luz como bacteriorhodopsina y halorhodopsin generan gradientes de iones bombeando iones desde y hacia la célula a través de la membrana plasmática. La energía almacenada en estos gradientes electroquímicos es luego convertida en ATP por la ATP sintasa. Este proceso es una forma de fotofosforilación. La capacidad de estas bombas accionadas por la luz para mover los iones a través de las membranas depende de los cambios impulsados por la luz en la estructura de un cofactor de retinol enterrado en el centro de la proteína.
Genética
Las arqueas usualmente tienen un solo cromosoma circular, con hasta 5,751,492 pares de bases en Methanosarcina acetivorans , el genoma de arquea más grande conocido. El pequeño genoma de 490.885 pares de bases de Nanoarchaeum equitans es una décima parte de este tamaño y se conoce el genoma archaeal más pequeño; se estima que contiene solo 537 genes que codifican proteínas. Piezas de ADN independientes más pequeñas, llamadas plásmidos , también se encuentran en arqueas. Los plásmidos se pueden transferir entre las células por contacto físico, en un proceso que puede ser similar a la conjugación bacteriana.

Las arqueas pueden ser infectadas por virus de ADN bicatenario que no están relacionados con ninguna otra forma de virus y tienen una variedad de formas inusuales, como botellas, varillas enganchadas o lágrimas. Estos virus han sido estudiados con mayor detalle en termófilos, particularmente los órdenes Sulfolobales y Thermoproteales. Recientemente se han aislado dos grupos de virus de ADN monocatenario que infectan arqueas. Un grupo está ejemplificado por el virus pleomórfico de Halorubrum 1 ("Pleolipoviridae") que infecta arqueas halófilas y el otro por el virus Aeropyrum en forma de espiral ("Spiraviridae") que infecta un huésped hipertermófilo (crecimiento óptimo a 90-95 ° C). Notablemente, este último virus tiene el mayor genoma de ssDNA actualmente informado. Las defensas contra estos virus pueden implicar interferencia de ARN a partir de secuencias repetitivas de ADN que están relacionadas con los genes de los virus.
Las arqueas son genéticamente distintas de las bacterias y eucariotas, con hasta un 15% de las proteínas codificadas por cualquier genoma de arquea que sea único para el dominio, aunque la mayoría de estos genes únicos no tienen una función conocida. Del resto de las proteínas únicas que tienen una función identificada, la mayoría pertenecen a Euryarchaea y están involucradas en la metanogénesis. Las proteínas que arqueas, bacterias y eucariotas comparten forman un núcleo común de funciones celulares, que se relaciona principalmente con la transcripción, la traducción y el metabolismo de nucleótidos. Otras características arqueológicas características son la organización de genes de función relacionada, como enzimas que catalizan pasos en la misma ruta metabólica hacia operones novedosos, y grandes diferencias en los genes de tRNA y sus aminoacil tRNA sintetasas.
La transcripción en arqueas se asemeja más a la transcripción eucariótica que a la bacteriana, con la ARN polimerasa arcaica muy cerca de su equivalente en eucariotas; mientras que la traducción archaeal muestra signos de equivalentes bacterianos y eukaryal. Aunque las arqueas solo tienen un tipo de ARN polimerasa, su estructura y función en la transcripción parece ser similar a la de la ARN polimerasa II eucariota, con ensamblajes de proteínas similares (factores de transcripción generales) que dirigen la unión de la ARN polimerasa al promotor de un gen , pero otros factores de transcripción archaeal están más cerca de los que se encuentran en las bacterias. La modificación postranscripcional es más simple que en los eucariotas, ya que la mayoría de los genes arqueos carecen de intrones, aunque hay muchos intrones en sus genes de ARN de transferencia y ARN ribosómico.
Transferencia de genes e intercambio genético
Halobacterium volcanii , una arquea halófila extrema, forma puentes citoplásmicos entre las células que parecen usarse para la transferencia de ADN de una célula a otra en cualquier dirección.
Cuando las arqueas hipertérmofílica Sulfolobus solfataricus y Sulfolobus acidocaldarius están expuestas a radiación UV que daña el ADN o a los agentes bleomicina o mitomicina C, se induce la agregación celular específica de la especie. La agregación en S. solfataricus no pudo ser inducida por otros factores estresantes físicos, como el cambio de pH o de temperatura, lo que sugiere que la agregación se induce específicamente por daño en el ADN. Ajon et al. mostró que la agregación celular inducida por UV media el intercambio de marcadores cromosómicos con alta frecuencia en S. acidocaldarius. Las tasas de recombinación excedieron las de cultivos no inducidos hasta en tres órdenes de magnitud. Frols et al. y Ajon et al. hipotetizó que la agregación celular mejora la transferencia de ADN específica de especie entre las células de Sulfolobus para proporcionar una reparación aumentada del ADN dañado por medio de recombinación homóloga. Esta respuesta puede ser una forma primitiva de interacción sexual similar a los sistemas de transformación bacteriana más estudiados que también están asociados con la transferencia de ADN específica de especie entre las células que conduce a la reparación homóloga de la recombinación del daño del ADN.
Reproducción
Las arqueas se reproducen asexualmente mediante fisión binaria o múltiple, fragmentación o gemación; la mitosis y la meiosis no ocurren, por lo tanto, si una especie de arquea existe en más de una forma, todas tienen el mismo material genético. La división celular se controla en un ciclo celular; después de que el cromosoma de la célula se replica y los dos cromosomas hijas se separan, la célula se divide. En el género Sulfolobus , el ciclo tiene características que son similares a los sistemas bacterianos y eucariotas. Los cromosomas se replican desde múltiples puntos de partida (orígenes de replicación) utilizando ADN polimerasas que se parecen a las enzimas eucariotas equivalentes.
En euryarchaea, la proteína de división celular FtsZ, que forma un anillo de contracción alrededor de la célula, y los componentes del tabique que se construyen en el centro de la célula, son similares a sus equivalentes bacterianos. En cren- y thaumarchaea, pero la maquinaria de división celular Cdv cumple un rol similar. Esta maquinaria está relacionada con la maquinaria eucariótica ESCRT-III que, si bien es mejor conocida por su papel en la clasificación celular, también se ha visto que cumple un papel en la separación entre células divididas, lo que sugiere un papel ancestral en la división celular.
Tanto las bacterias como los eucariotas, pero no las arqueas, producen esporas. Algunas especies de Haloarchaea experimentan cambios fenotípicos y crecen en diferentes tipos de células, incluyendo estructuras de pared gruesa que son resistentes al choque osmótico y permiten que las arqueas sobrevivan en el agua a bajas concentraciones de sal, pero estas no son estructuras reproductivas y pueden ayudarlas. alcanzar nuevos hábitats
Ecología
Hábitats

Las arqueas existen en una amplia gama de hábitats, y como una parte importante de los ecosistemas mundiales, pueden representar aproximadamente el 20% de las células microbianas en los océanos. Los primeros arqueos descubiertos fueron extremófilos. De hecho, algunas arqueas sobreviven a altas temperaturas, a menudo por encima de 100 ° C (212 ° F), como se encuentra en los géiseres, fumadores negros y pozos de petróleo. Otros hábitats comunes incluyen hábitats muy fríos y agua altamente salina, ácida o alcalina, pero las arqueas incluyen mesófilos que crecen en condiciones suaves, en pantanos y marismas, aguas residuales, océanos, tracto intestinal de animales y suelos.
Las arqueas extremófilas son miembros de cuatro grupos fisiológicos principales. Estos son los halófilos, termófilos, alcalófilos y acidófilos. Estos grupos no son exhaustivos o específicos del filo, ni son mutuamente excluyentes, ya que algunas arqueas pertenecen a varios grupos. No obstante, son un punto de partida útil para la clasificación.
Los halófilos, incluido el género Halobacterium , viven en entornos extremadamente salinos, como los lagos salados, y superan en número a sus homólogos bacterianos en salinidades superiores al 20-25%. Los termófilos crecen mejor a temperaturas superiores a 45 ° C (113 ° F), en lugares tales como aguas termales; Las arqueas hipertermofílicas crecen de manera óptima a temperaturas superiores a 80 ° C (176 ° F). La cepa 116 de Archaeal Methanopyrus kandleri puede incluso reproducirse a 122 ° C (252 ° F), la temperatura más alta registrada de cualquier organismo.
Existen otras arqueas en condiciones muy ácidas o alcalinas. Por ejemplo, uno de los acidófilos arcaicos más extremos es Picrophilus torridus , que crece a pH 0, que es equivalente a prosperar en ácido sulfúrico 1.2 molar.
Esta resistencia a los ambientes extremos ha convertido a las arqueas en el foco de la especulación sobre las posibles propiedades de la vida extraterrestre. Algunos hábitats extremófilos no son muy diferentes a los de Marte, lo que sugiere que los microbios viables podrían ser transferidos entre los planetas en los meteoritos.
Recientemente, varios estudios han demostrado que las arqueas existen no solo en ambientes mesofílicos y termófilos, sino que también están presentes, a veces en grandes cantidades, a bajas temperaturas también. Por ejemplo, las arqueas son comunes en ambientes oceánicos fríos como los mares polares. Aún más significativo es el gran número de arqueas que se encuentran en los océanos del mundo en hábitats no extremos entre la comunidad de plancton (como parte del picoplancton). Aunque estas arqueas pueden estar presentes en cantidades extremadamente altas (hasta 40% de la biomasa microbiana), casi ninguna de estas especies ha sido aislada y estudiada en cultivo puro. En consecuencia, nuestra comprensión del papel de las arqueas en la ecología de los océanos es rudimentaria, por lo que su influencia total en los ciclos biogeoquímicos globales permanece en gran parte inexplorada. Algunos Crenarchaeota marinos son capaces de nitrificación, sugiriendo que estos organismos pueden afectar el ciclo del nitrógeno oceánico, aunque estos Crenarchaeota oceánicos también pueden usar otras fuentes de energía. Enormes cantidades de arqueas también se encuentran en los sedimentos que cubren el fondo del mar, con estos organismos que constituyen la mayoría de las células vivas a profundidades de más de 1 metro por debajo del fondo oceánico. Se ha demostrado que en todos los sedimentos superficiales oceánicos (de 1000 a 10 000 m de profundidad del agua), el impacto de la infección viral es mayor en las arqueas que en las bacterias y la lisis inducida por virus de las cuentas de arqueas hasta en un tercio del total de biomasa microbiana muerta, lo que resulta en la liberación de ~ 0.3 a 0.5 gigatoneladas de carbono por año en todo el mundo. Enormes cantidades de arqueas también se encuentran en los sedimentos que cubren el fondo del mar, con estos organismos que constituyen la mayoría de las células vivas a profundidades de más de 1 metro por debajo del fondo oceánico. Se ha demostrado que en todos los sedimentos superficiales oceánicos (de 1000 a 10 000 m de profundidad del agua), el impacto de la infección viral es mayor en las arqueas que en las bacterias y la lisis inducida por virus de las cuentas de arqueas hasta en un tercio del total de biomasa microbiana muerta, lo que resulta en la liberación de ~ 0.3 a 0.5 gigatoneladas de carbono por año en todo el mundo. Enormes cantidades de arqueas también se encuentran en los sedimentos que cubren el fondo del mar, con estos organismos que constituyen la mayoría de las células vivas a profundidades de más de 1 metro por debajo del fondo oceánico. Se ha demostrado que en todos los sedimentos superficiales oceánicos (de 1000 a 10 000 m de profundidad del agua), el impacto de la infección viral es mayor en las arqueas que en las bacterias y la lisis inducida por virus de las cuentas de arqueas hasta en un tercio del total de biomasa microbiana muerta, lo que resulta en la liberación de ~ 0.3 a 0.5 gigatoneladas de carbono por año en todo el mundo.
Papel en el ciclismo químico
Archaea recicla elementos como el carbono, el nitrógeno y el azufre a través de sus diversos hábitats. Aunque estas actividades son vitales para la función normal del ecosistema, las arqueas también pueden contribuir a los cambios producidos por los seres humanos e incluso causar contaminación.
Archaea lleva a cabo muchos pasos en el ciclo del nitrógeno. Esto incluye tanto las reacciones que eliminan el nitrógeno de los ecosistemas (como la respiración y desnitrificación basadas en nitratos) como los procesos que introducen nitrógeno (como la asimilación de nitratos y la fijación de nitrógeno). Los investigadores descubrieron recientemente la participación de arqueas en las reacciones de oxidación de amoníaco. Estas reacciones son particularmente importantes en los océanos. Las arqueas también parecen ser cruciales para la oxidación del amoníaco en los suelos. Producen nitrito, que otros microbios luego oxidan a nitrato. Las plantas y otros organismos consumen este último.
En el ciclo de azufre, las arqueas que crecen oxidando compuestos de azufre liberan este elemento de las rocas, haciéndolo disponible para otros organismos, pero las arqueas que lo hacen, como Sulfolobus , producen ácido sulfúrico como producto de desecho y el crecimiento de estos organismos en minas abandonadas puede contribuir al drenaje ácido de la mina y a otros daños ambientales.
En el ciclo del carbono, el metanogen archaea elimina el hidrógeno y juega un papel importante en la descomposición de la materia orgánica por parte de las poblaciones de microorganismos que actúan como descomponedores en los ecosistemas anaeróbicos, como los sedimentos, las marismas y las obras de tratamiento de aguas residuales.
Interacciones con otros organismos

Las interacciones bien caracterizadas entre las arqueas y otros organismos son mutuas o comensales. No hay ejemplos claros de patógenos o parásitos archaeal conocidos, pero se ha sugerido que algunas especies de metanógenos están involucradas en infecciones en la boca, y Nanoarchaeum equitans puede ser un parásito de otra especie de arqueas, ya que solo sobrevive y se reproduce dentro del células del Crenarchaeon Ignicoccus hospitalis , y parece no ofrecer ningún beneficio para su huésped. Las conexiones entre las células archaeas también se pueden encontrar entre Archaeal Richmond Mine Acidophilic Nanoorganisms (ARMAN) y otra especie de archaea llamada Thermoplasmatales, dentro de las biopelículas de drenaje ácido de la mina. Aunque la naturaleza de esta relación es desconocida, es distinta de la de Nanarchaeaum-Ignicoccus en que las células ARMAN ultracortas son generalmente independientes de las células de Thermoplasmatales.
Mutualismo
Un ejemplo bien entendido de mutualismo es la interacción entre los protozoos y las arqueas metanogénicas en el tracto digestivo de los animales que digieren la celulosa, como los rumiantes y las termitas. En estos ambientes anaeróbicos, los protozoos descomponen la celulosa vegetal para obtener energía. Este proceso libera hidrógeno como un producto de desecho, pero los altos niveles de hidrógeno reducen la producción de energía. Cuando los metanógenos convierten el hidrógeno en metano, los protozoos se benefician con más energía.
En los protozoos anaerobios, como Plagiopyla frontata , las arqueas residen dentro de los protozoos y consumen hidrógeno producido en sus hidrogenosomas. Archaea también se asocia con organismos más grandes. Por ejemplo, el simbionte Cenarchaeum marino de las arqueas vive dentro (es un endosimbionte de) la esponja Axinella mexicana .
Comensalismo
Archaea también puede ser comensales, beneficiándose de una asociación sin ayudar o dañar al otro organismo. Por ejemplo, el metanogeno Methanobrevibacter smithii es con mucho el archaean mas comun en la flora humana, que compone aproximadamente uno de cada diez de todos los procariotas en el intestino humano. En termitas y en humanos, estos metanógenos pueden de hecho ser mutualistas, interactuando con otros microbios en el intestino para ayudar a la digestión. Las comunidades de Archaean también se asocian con una variedad de otros organismos, como en la superficie de los corales y en la región del suelo que rodea las raíces de las plantas (la rizosfera).
Importancia en tecnología e industria
Las arqueas extremófilas, particularmente aquellas resistentes al calor oa los extremos de acidez y alcalinidad, son una fuente de enzimas que funcionan en estas duras condiciones. Estas enzimas han encontrado muchos usos. Por ejemplo, las ADN polimerasas termoestables, como la Pfu ADN polimerasa de Pyrococcus furiosus , revolucionaron la biología molecular permitiendo que la reacción en cadena de la polimerasa se usara en la investigación como una técnica simple y rápida para clonar ADN. En la industria, amilasas, galactosidasas y pululanasas en otras especies de Pyrococcus esa función a más de 100 ° C (212 ° F) permite el procesamiento de alimentos a altas temperaturas, como la producción de leche baja en lactosa y suero de leche. Las enzimas de estas arqueas termófilas también tienden a ser muy estables en solventes orgánicos, permitiendo su uso en procesos ecológicos en la química verde que sintetizan compuestos orgánicos. Esta estabilidad los hace más fáciles de usar en biología estructural. En consecuencia, las contrapartes de enzimas bacterianas o eucarióticas de arqueas extremófilas se utilizan a menudo en estudios estructurales.
En contraste con el rango de aplicaciones de las enzimas arqueanas, el uso de los propios organismos en biotecnología está menos desarrollado. Las arqueas metanogénicas son una parte vital del tratamiento de aguas residuales, ya que son parte de la comunidad de microorganismos que llevan a cabo la digestión anaeróbica y producen biogás. En el procesamiento de minerales, las arqueas acidófilas son prometedoras para la extracción de metales de los minerales, incluidos el oro, el cobalto y el cobre.
Archaea alberga una nueva clase de antibióticos potencialmente útiles. Algunas de estas archaeocins se han caracterizado, pero se cree que existen cientos más, especialmente dentro de Haloarchaea y Sulfolobus . Estos compuestos difieren en su estructura de los antibióticos bacterianos, por lo que pueden tener nuevos modos de acción. Además, pueden permitir la creación de nuevos marcadores seleccionables para su uso en biología molecular archaeal.
Obtenido de: https://en.wikipedia.org/wiki/Archaea