Bacteria
Definición
| Gama de Bacteria Temporal: Archean o anterior - presente Had'n Archean Proterozoico Pha. | |
|---|---|
 | |
| Microfotografía electrónica de escaneo de varillas de Escherichia coli | |
| clasificación cientifica | |
| Dominio: | Bacteria Woese, Kandler y Wheelis, 1990 |
| Phyla | |
Acidobacteria Actinobacteria Aquificae Armatimonadetes Bacteroidetes Caldiserica Chlamydiae Chlorobi Chloroflexi Chrysiogenetes Cyanobacteria Deferribacteres Deinococcus-Thermus Dictyoglomi Elusimicrobia Fibrobacteria Firmicutes Fusobacteria Gemmatimonadetes Lentisphaerae Nitrospirae Planctomycetes Proteobacteria Espiroquetas Sinergistas Tenericutes Termodesulfobacteria Thermotogae Verrucomicrobia | |
| Sinónimos | |
Eubacteria Woese & Fox, 1977 | |
Normalmente hay 40 millones de células bacterianas en un gramo de suelo y un millón de células bacterianas en un mililitro de agua dulce. Hay aproximadamente 5 × 10 bacterias en la Tierra, formando una biomasa que excede la de todas las plantas y animales. Las bacterias son vitales en muchas etapas del ciclo de nutrientes reciclando nutrientes como la fijación de nitrógeno de la atmósfera. El ciclo de nutrientes incluye la descomposición de los cadáveres y las bacterias son responsables de la etapa de putrefacción en este proceso. En las comunidades biológicas que rodean los respiraderos hidrotermales y las filtraciones frías, las bacterias extremófilas proporcionan los nutrientes necesarios para mantener la vida mediante la conversión de compuestos disueltos, como el sulfuro de hidrógeno y el metano, en energía. En marzo de 2013, se publicaron los datos informados por los investigadores en octubre de 2012. Se sugirió que las bacterias prosperan en la Fosa de las Marianas, que con una profundidad de hasta 11 kilómetros es la parte más profunda conocida de los océanos. Otros investigadores informaron estudios relacionados que los microbios crecen dentro de las rocas hasta 580 metros debajo del fondo del mar a 2,6 kilómetros de océano frente a la costa del noroeste de los Estados Unidos. Según uno de los investigadores, "se pueden encontrar microbios en todas partes, son extremadamente adaptable a las condiciones, y sobrevivir donde sea que estén ".
La famosa noción de que las células bacterianas en el cuerpo humano superan a las células humanas en un factor de 10: 1 ha sido desacreditada. Hay aproximadamente 39 billones de células bacterianas en la microbiota humana personificadas por una "referencia" de 70 kg de machos de 170 cm de altura, mientras que hay 30 billones de células humanas en el cuerpo. Esto significa que, aunque tienen la ventaja en números reales, solo es un 30% y no un 900%.
El número más grande existe en la flora intestinal y un gran número en la piel. La gran mayoría de las bacterias en el cuerpo se vuelven inofensivas por los efectos protectores del sistema inmune, aunque muchas son beneficiosas, particularmente en la flora intestinal. Sin embargo, varias especies de bacterias son patógenas y causan enfermedades infecciosas, como el cólera, la sífilis, el ántrax, la lepra y la peste bubónica. Las enfermedades bacterianas fatales más comunes son las infecciones respiratorias, con la tuberculosis que mata a aproximadamente 2 millones de personas por año, principalmente en el África subsahariana. En los países desarrollados, los antibióticos se usan para tratar infecciones bacterianas y también se usan en la agricultura, lo que hace que la resistencia a los antibióticos sea un problema creciente. En la industria, las bacterias son importantes en el tratamiento de aguas residuales y la descomposición de los derrames de petróleo,
Una vez consideradas como plantas que constituyen la clase Schizomycetes , las bacterias ahora se clasifican como procariotas. A diferencia de las células de animales y otros eucariotas, las células bacterianas no contienen un núcleo y raramente albergan organelos unidos a la membrana. Aunque el término bacterias tradicionalmente incluía todos los procariotas, la clasificación científica cambió después del descubrimiento en la década de 1990 de que los procariotas consisten en dos grupos muy diferentes de organismos que evolucionaron a partir de un ancestro común antiguo. Estos dominios evolutivos se llaman Bacteria y Archaea .
Etimología
Origen y evolución tempranaLa palabra bacteria es el plural de la Nueva bacteria latina , que es la latinización del griego βακτήριον ( bakterion ), el diminutivo de βακτηρία ( bakteria ), que significa "bastón, bastón", porque los primeros que se descubrieron fueron en forma de barra
Los antepasados de las bacterias modernas eran microorganismos unicelulares que fueron las primeras formas de vida que aparecieron en la Tierra, hace unos 4.000 millones de años. Durante aproximadamente 3 mil millones de años, la mayoría de los organismos eran microscópicos, y las bacterias y arqueas eran las formas de vida dominantes. Aunque existen fósiles bacterianos, como los estromatolitos, su falta de morfología distintiva impide que se utilicen para examinar la historia de la evolución bacteriana, o hasta la fecha del origen de una especie bacteriana particular. Sin embargo, las secuencias de genes se pueden usar para reconstruir la filogenia bacteriana, y estos estudios indican que las bacterias divergieron primero del linaje archaal / eucariótico. El ancestro común más reciente de bacterias y arqueas fue probablemente un hipertermófilo que vivió hace unos 2.500 a 3.200 millones de años.
Las bacterias también estuvieron involucradas en la segunda gran divergencia evolutiva, la de las arqueas y las eucariotas. Aquí, los eucariotas resultaron de la entrada de bacterias antiguas en asociaciones endosimbióticas con los antepasados de células eucariotas, que posiblemente estaban relacionadas con las arqueas. Esto implicó la absorción por células proto-eucariotas de alfaproteobacterialsymbionts para formar mitocondrias o hidrogenosomas, que todavía se encuentran en todos los conocidos Eukarya (a veces en forma muy reducida, por ejemplo, en antiguos protozoos "amitocondriales"). Más tarde, algunos eucariotas que ya contenían mitocondrias también engullían organismos similares a las cianobacterias, lo que llevó a la formación de cloroplastos en algas y plantas. Esto se conoce como endosimbiosis secundaria.
Morfología

Las bacterias muestran una gran diversidad de formas y tamaños, llamadas morfologías. Las células bacterianas son aproximadamente un décimo del tamaño de las células eucarióticas y típicamente tienen una longitud de 0,5-5,0 micrómetros. Sin embargo, algunas especies son visibles a simple vista, por ejemplo, Thiomargarita namibiensis mide hasta medio milímetro y Epulopiscium fishelsoni alcanza los 0.7 mm. Entre las bacterias más pequeñas se encuentran miembros del género Mycoplasma, que mide solo 0.3 micrómetros, tan pequeños como los virus más grandes. Algunas bacterias pueden ser incluso más pequeñas, pero estas ultramicrobacterias no están bien estudiadas.
La mayoría de las especies bacterianas son esféricas, llamadas cocos ( cantar coccus, del griego kókkos , grano, semilla), o en forma de vara, llamados bacilos ( sing . Bacillus, del latín baculus , stick). Algunas bacterias, llamadas vibrio , tienen forma de varillas ligeramente curvadas o en forma de coma; otras pueden tener forma de espiral, llamadas espirillas , o bien enrolladas, llamadas espiroquetas.. Se ha descrito un pequeño número de otras formas inusuales, como las bacterias en forma de estrella. Esta amplia variedad de formas está determinada por la pared celular bacteriana y el citoesqueleto, y es importante porque puede influir en la capacidad de las bacterias para adquirir nutrientes, adherirse a las superficies, nadar a través de los líquidos y escapar de los depredadores.

Muchas especies bacterianas existen simplemente como células individuales, otras se asocian en patrones característicos: neisseria forman diploides (pares), Streptococcus forman cadenas y Staphylococcus agrupan en racimos de "racimo de uvas". Las bacterias también pueden agruparse para formar estructuras multicelulares más grandes, como los filamentos alargados de Actinobacteria , los agregados de Myxobacteria y las hifas complejas de Streptomyces.. Estas estructuras multicelulares a menudo solo se ven en ciertas condiciones. Por ejemplo, cuando carecen de aminoácidos, las mixobacterias detectan las células circundantes en un proceso conocido como detección de quórum, migran entre sí y se agregan para formar cuerpos fructíferos de hasta 500 micrómetros de largo y que contienen aproximadamente 100.000 células bacterianas. En estos cuerpos fructíferos, las bacterias realizan tareas separadas; por ejemplo, aproximadamente una de cada diez células migran a la parte superior de un cuerpo fructífero y se diferencian en un estado durmiente especializado llamado mixospora, que es más resistente al secado y a otras condiciones ambientales adversas.
Las bacterias a menudo se adhieren a las superficies y forman agregaciones densas llamadas biofilms, y formaciones más grandes conocidas como esteras microbianas. Estos biofilms y tapetes pueden variar desde unos pocos micrómetros de grosor hasta hasta medio metro de profundidad, y pueden contener múltiples especies de bacterias, protistas y arqueas. Las bacterias que viven en biofilms muestran una disposición compleja de células y componentes extracelulares, formando estructuras secundarias, como microcolonias, a través de las cuales hay redes de canales que permiten una mejor difusión de nutrientes. En ambientes naturales, como el suelo o las superficies de las plantas, la mayoría de las bacterias están unidas a las superficies de las biopelículas. Las biopelículas también son importantes en medicina, ya que estas estructuras a menudo están presentes durante infecciones bacterianas crónicas o en infecciones de dispositivos médicos implantados,
Estructura celular

Estructuras intracelulares
La célula bacteriana está rodeada por una membrana celular que está compuesta principalmente por fosfolípidos. Esta membrana encierra el contenido de la célula y actúa como una barrera para contener nutrientes, proteínas y otros componentes esenciales del citoplasma dentro de la célula. A diferencia de las células eucariotas, las bacterias generalmente carecen de grandes estructuras unidas a la membrana en su citoplasma, como un núcleo, mitocondrias, cloroplastos y otros orgánulos presentes en las células eucarióticas. Sin embargo, algunas bacterias tienen organelos unidos a proteínas en el citoplasma que compartimentan aspectos del metabolismo bacteriano, como el carboxisoma. Además, las bacterias tienen un citoesqueleto multicomponente para controlar la localización de proteínas y ácidos nucleicos dentro de la célula, y para administrar el proceso de división celular.
Muchas reacciones bioquímicas importantes, como la generación de energía, ocurren debido a los gradientes de concentración a través de las membranas, creando una diferencia potencial similar a la de una batería. La falta general de membranas internas en las bacterias significa que estas reacciones, como el transporte de electrones, ocurren a través de la membrana celular entre el citoplasma y el exterior de la célula o periplasma. Sin embargo, en muchas bacterias fotosintéticas, la membrana plasmática está muy plegada y llena la mayor parte de la célula con capas de membrana captadora de luz. Estos complejos de recolección de luz incluso pueden formar estructuras encerradas en lípidos llamadas clorosomas en bacterias verdes de azufre.
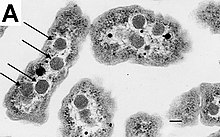
La mayoría de las bacterias no tienen un núcleo unido a la membrana, y su material genético es típicamente un único cromosoma bacteriano circular de ADN ubicado en el citoplasma en un cuerpo de forma irregular llamado nucleoide. El nucleoide contiene el cromosoma con sus proteínas y ARN asociados. Al igual que todos los organismos vivos, las bacterias contienen ribosomas para la producción de proteínas, pero la estructura del ribosoma bacteriano es diferente de la de los eucariotas y Archaea.
Algunas bacterias producen gránulos de almacenamiento de nutrientes intracelulares, como glucógeno, polifosfato, azufre o polihidroxialcanoatos. Ciertas especies bacterianas, como las cianobacterias fotosintéticas, producen vacuolas de gas internas que utilizan para regular su flotabilidad, lo que les permite moverse hacia arriba o hacia abajo en capas de agua con diferentes intensidades de luz y niveles de nutrientes.
Estructuras extracelulares
Alrededor de la parte exterior de la membrana celular está la pared celular. Las paredes de las células bacterianas están hechas de peptidoglicano (llamado "mureína" en fuentes más antiguas), que está hecho de cadenas de polisacáridos reticuladas por péptidos que contienen D-aminoácidos. Las paredes de las células bacterianas son diferentes de las paredes celulares de las plantas y los hongos, que están hechos de celulosa y quitina, respectivamente. La pared celular de las bacterias también es distinta de la de Archaea, que no contiene peptidoglicano. La pared celular es esencial para la supervivencia de muchas bacterias, y el antibiótico penicilina es capaz de matar bacterias al inhibir un paso en la síntesis de peptidoglicano.
En términos generales, existen dos tipos diferentes de pared celular en bacterias, llamadas Gram-positivas y Gram-negativas. Los nombres provienen de la reacción de las células a la tinción de Gram, una prueba de larga data para la clasificación de especies bacterianas.
Las bacterias Gram-positivas poseen una pared celular gruesa que contiene muchas capas de peptidoglicanos y ácidos teicoicos. Por el contrario, las bacterias Gram-negativas tienen una pared celular relativamente delgada que consiste en unas pocas capas de peptidoglicano rodeadas por una segunda membrana lipídica que contiene lipopolisacáridos y lipoproteínas. La mayoría de las bacterias tienen la pared de células Gram-negativas, y solo las Actinobacterias Firmicutesand (anteriormente conocidas como bacterias Gram-positivas bajas G + C y altas G + C, respectivamente) tienen la disposición Gram-positiva alternativa. Estas diferencias en la estructura pueden producir diferencias en la susceptibilidad a los antibióticos; por ejemplo, la vancomicina puede matar solo bacterias Gram-positivas y es ineficaz contra patógenos Gram-negativos, como Haemophilus influenzae o Pseudomonas aeruginosa. Algunas bacterias tienen estructuras de pared celular que no son ni clásicamente Gram-positivas ni Gram-negativas. Esto incluye bacterias clínicamente importantes como las micobacterias que tienen una pared gruesa de células de peptidoglucano como una bacteria Gram-positiva, pero también una segunda capa externa de lípidos.
En muchas bacterias, una capa S de moléculas proteicas rígidamente dispuestas cubre el exterior de la célula. Esta capa proporciona protección química y física para la superficie de la célula y puede actuar como una barrera de difusión macromolecular. Las capas S tienen funciones diversas pero en su mayoría poco conocidas, pero se sabe que actúan como factores de virulencia en Campylobacter y contienen enzimas superficiales en Bacillus stearothermophilus .
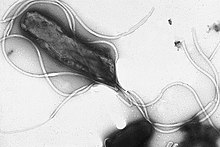
Los flagelos son estructuras de proteínas rígidas, de unos 20 nanómetros de diámetro y hasta 20 micras de longitud, que se utilizan para la motilidad. Los flagelos son impulsados por la energía liberada por la transferencia de iones a través de un gradiente electroquímico a través de la membrana celular.
Las fimbrias (a veces llamadas "pili de unión") son filamentos finos de proteína, usualmente 2-10 nanómetros de diámetro y hasta varios micrómetros de longitud. Se distribuyen sobre la superficie de la célula y se asemejan a pelos finos cuando se ven bajo el microscopio electrónico. Se cree que las fimbrias están involucradas en la fijación a superficies sólidas o a otras células, y son esenciales para la virulencia de algunos patógenos bacterianos. Pili ( sing . Pilus) son apéndices celulares, ligeramente más grandes que las fimbrias, que pueden transferir material genético entre las células bacterianas en un proceso llamado conjugación, donde se les llama conjugación pili o pili sexual (ver genética bacteriana, abajo). También pueden generar movimiento donde se llaman pili tipo IV.
Glycocalyx es producido por muchas bacterias para rodear sus células, y varía en complejidad estructural: desde una capa de lodo desorganizado de sustancias poliméricas extracelulares a una cápsula altamente estructurada. Estas estructuras pueden proteger a las células del engullimiento de las células eucariotas, como los macrófagos (parte del sistema inmune humano). También pueden actuar como antígenos y participar en el reconocimiento celular, así como ayudar a la fijación a las superficies y la formación de biofilms.
El ensamblaje de estas estructuras extracelulares depende de los sistemas de secreción bacteriana. Estas transfieren proteínas del citoplasma al periplasma o al entorno de la célula. Se conocen muchos tipos de sistemas de secreción y estas estructuras a menudo son esenciales para la virulencia de los patógenos, por lo que se estudian intensamente.
Endosporas
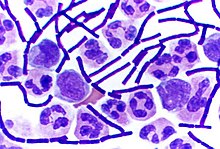
Ciertos géneros de bacterias Gram-positivas, como Bacillus , Clostridium , Sporohalobacter , Anaerobacter y Heliobacterium, pueden formar estructuras latentes altamente resistentes llamadas endosporas . Las endosporas se desarrollan dentro del citoplasma de la célula; en general, se desarrolla una única endospora en cada célula. Cada endospora contiene un núcleo de ADN y ribosomas rodeados por una capa de corteza y protegidos por una capa rígida multicapa compuesta de peptidoglicano y una variedad de proteínas.
Las endosporas no muestran metabolismo detectable y pueden resistir tensiones físicas y químicas extremas, como niveles elevados de luz ultravioleta, radiación gamma, detergentes, desinfectantes, calor, congelación, presión y desecación. En este estado latente, estos organismos pueden seguir siendo viables durante millones de años, y las endosporas incluso permiten que las bacterias sobrevivan a la exposición al vacío y a la radiación en el espacio. Las bacterias formadoras de endosporas también pueden causar enfermedades: por ejemplo, el ántrax puede contraerse mediante la inhalación de endosporas de Bacillus anthracis , y la contaminación de las heridas profundas por punción con endosporas de Clostridium tetanicausa el tétanos.
Metabolismo
Las bacterias exhiben una variedad extremadamente amplia de tipos metabólicos. La distribución de los rasgos metabólicos dentro de un grupo de bacterias se ha utilizado tradicionalmente para definir su taxonomía, pero estos rasgos a menudo no se corresponden con las clasificaciones genéticas modernas. El metabolismo bacteriano se clasifica en grupos nutricionales sobre la base de tres criterios principales: la fuente de energía, los donantes de electrones utilizados y la fuente de carbono utilizada para el crecimiento.
Las bacterias obtienen energía de la luz mediante la fotosíntesis (llamada fototrofía) o descomponen los compuestos químicos mediante la oxidación (llamada quimiotrofía). Los químicos quimiotróficos utilizan compuestos químicos como fuente de energía mediante la transferencia de electrones de un donador de electrones dado a un aceptor de electrones terminal en una reacción redox. Esta reacción libera energía que puede usarse para impulsar el metabolismo. Los quimiotrofos se dividen además por los tipos de compuestos que utilizan para transferir electrones. Las bacterias que usan compuestos inorgánicos como hidrogren, monóxido de carbono o amoniaco como fuentes de electrones se llaman litotrofos, mientras que las que usan compuestos orgánicos se llaman organótrofos. Los compuestos usados para recibir electrones también se usan para clasificar bacterias: los organismos aeróbicos usan oxígeno como el aceptor de electrones terminal,
Muchas bacterias obtienen su carbono de otro carbono orgánico, llamado heterotrofia. Otros, como las cianobacterias y algunas bacterias moradas, son autótrofos, lo que significa que obtienen carbono celular mediante la fijación del dióxido de carbono. En circunstancias inusuales, el gas metano puede ser utilizado por bacterias metanotróficas como fuente de electrones y como sustrato para el anabolismo de carbono.
| Tipo nutricional | Fuente de energía | Fuente de carbono | Ejemplos |
|---|---|---|---|
| Fototrofos | Luz de sol | Compuestos orgánicos (fotoheterótrofos) o fijación de carbono (fotoautótrofos) | Cianobacterias, bacterias de azufre verde, Chloroflexi o bacterias púrpuras |
| Lithotrophs | Compuestos inorgánicos | Compuestos orgánicos (lithoheterotrophs) o fijación de carbón (lithoautotrophs) | Thermodesulfobacteria, Hydrogenophilaceae o Nitrospirae |
| Organotrofos | Compuestos orgánicos | Compuestos orgánicos (quimioheterótrofos) o fijación de carbono (quimioautótrofos) | Bacillus , Clostridium o Enterobacteriaceae |
En muchos sentidos, el metabolismo bacteriano proporciona rasgos que son útiles para la estabilidad ecológica y para la sociedad humana. Un ejemplo es que algunas bacterias tienen la capacidad de fijar nitrógeno gaseoso utilizando la enzima nitrogenasa. Este rasgo ambientalmente importante se puede encontrar en bacterias de la mayoría de los tipos metabólicos enumerados anteriormente. Esto conduce a los procesos ecológicamente importantes de desnitrificación, reducción de sulfato y acetogénesis, respectivamente. Los procesos metabólicos bacterianos también son importantes en las respuestas biológicas a la contaminación; por ejemplo, las bacterias reductoras de sulfato son en gran parte responsables de la producción de las formas altamente tóxicas de mercurio (metil- y dimetilmercurio) en el medio ambiente. Los anaerobios no respiratorios usan la fermentación para generar energía y reducir la potencia, secretando subproductos metabólicos (como el etanol en la elaboración de la cerveza) como desecho.
Crecimiento y reproducción
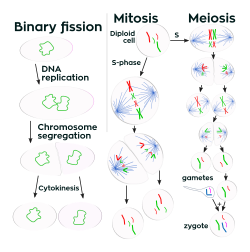
A diferencia de los organismos multicelulares, los aumentos en el tamaño celular (crecimiento celular) y la reproducción por división celular están estrechamente relacionados en organismos unicelulares. Las bacterias crecen a un tamaño fijo y luego se reproducen a través de la fisión binaria, una forma de reproducción asexual. En condiciones óptimas, las bacterias pueden crecer y dividirse extremadamente rápido, y las poblaciones bacterianas pueden duplicarse tan rápido como cada 9.8 minutos. En la división celular, se producen dos células idénticas de hija clonada. Algunas bacterias, aunque se reproducen asexualmente, forman estructuras reproductivas más complejas que ayudan a dispersar las células hijas recién formadas. Los ejemplos incluyen la formación de cuerpos fructíferos por Myxobacteria y la formación de hifas aéreas por Streptomyces, o en ciernes. El brote involucra una célula que forma una protuberancia que se desprende y produce una célula hija.

En el laboratorio, las bacterias generalmente se cultivan utilizando medios sólidos o líquidos. Los medios de crecimiento sólidos, como las placas de agar, se usan para aislar cultivos puros de una cepa bacteriana. Sin embargo, los medios de crecimiento líquido se usan cuando se requieren medidas de crecimiento o grandes volúmenes de células. El crecimiento en medios líquidos agitados se produce como una suspensión celular uniforme, haciendo que los cultivos sean fáciles de dividir y transferir, aunque es difícil aislar las bacterias individuales de los medios líquidos. El uso de medios selectivos (medios con nutrientes específicos agregados o deficientes, o con antibióticos añadidos) puede ayudar a identificar organismos específicos.
La mayoría de las técnicas de laboratorio para el crecimiento de bacterias utilizan altos niveles de nutrientes para producir grandes cantidades de células de manera económica y rápida. Sin embargo, en ambientes naturales, los nutrientes son limitados, lo que significa que las bacterias no pueden continuar reproduciéndose indefinidamente. Esta limitación de nutrientes ha conducido a la evolución de diferentes estrategias de crecimiento (ver teoría de selección r / K). Algunos organismos pueden crecer extremadamente rápido cuando los nutrientes están disponibles, como la formación de floraciones de algas (y cianobacterias) que a menudo ocurren en los lagos durante el verano. Otros organismos tienen adaptaciones a entornos hostiles, como la producción de múltiples antibióticos por Streptomyces que inhiben el crecimiento de microorganismos competidores. En la naturaleza, muchos organismos viven en comunidades (por ejemplo, biofilms) que pueden permitir un mayor suministro de nutrientes y protección contra el estrés ambiental. Estas relaciones pueden ser esenciales para el crecimiento de un organismo o grupo de organismos en particular (sintrofia).
El crecimiento bacteriano sigue cuatro fases. Cuando una población de bacterias ingresa primero a un ambiente con alto contenido de nutrientes que permite el crecimiento, las células necesitan adaptarse a su nuevo entorno. La primera fase de crecimiento es la fase de retraso, un período de crecimiento lento cuando las células se adaptan al entorno de alto contenido de nutrientes y se preparan para un crecimiento rápido. La fase de latencia tiene altas tasas de biosíntesis, ya que se producen las proteínas necesarias para un crecimiento rápido. La segunda fase de crecimiento es la fase logarítmica, también conocida como fase exponencial. La fase de registro está marcada por un rápido crecimiento exponencial. La velocidad a la que las células crecen durante esta fase se conoce como la tasa de crecimiento ( k ), y el tiempo que tardan las células en duplicarse se conoce como el tiempo de generación ( g).) Durante la fase de registro, los nutrientes se metabolizan a velocidad máxima hasta que uno de los nutrientes se agota y comienza a limitar el crecimiento. La tercera fase de crecimiento es la fase estacionaria y es causada por nutrientes agotados. Las células reducen su actividad metabólica y consumen proteínas celulares no esenciales. La fase estacionaria es una transición del crecimiento rápido al estado de respuesta al estrés y existe una mayor expresión de genes implicados en la reparación del ADN, el metabolismo antioxidante y el transporte de nutrientes. La fase final es la fase de muerte donde las bacterias se quedan sin nutrientes y mueren.
Genética
La mayoría de las bacterias tienen un solo cromosoma circular que puede variar en tamaño desde solo 160,000 pares de bases en la bacteria endosimbiótica Carsonella ruddii , hasta 12,200,000 pares de bases (12.2 Mbp) en la bacteria Sorangium cellulosum que vive en el suelo . Hay muchas excepciones a esto, por ejemplo, algunas especies de Streptomyces y Borrelia contienen un solo cromosoma lineal, mientras que algunas especies de Vibrio contienen más de un cromosoma. Las bacterias también pueden contener plásmidos, pequeños ADN extracromosómicos que pueden contener genes para diversas funciones útiles, como resistencia a los antibióticos, capacidades metabólicas o diversos factores de virulencia.
Los genomas de bacterias generalmente codifican unos pocos cientos a unos miles de genes. Los genes en los genomas bacterianos suelen ser un único tramo continuo de ADN y aunque existen varios tipos diferentes de intrones en las bacterias, estos son mucho más raros que en los eucariotas.
Las bacterias, como organismos asexuales, heredan una copia idéntica de los genomas de los padres y son clonales. Sin embargo, todas las bacterias pueden evolucionar mediante la selección de cambios en su ADN de material genético causado por recombinación genética o mutaciones. Las mutaciones provienen de errores cometidos durante la replicación del ADN o de la exposición a mutágenos. Las tasas de mutación varían ampliamente entre diferentes especies de bacterias e incluso entre diferentes clones de una sola especie de bacteria. Los cambios genéticos en los genomas bacterianos provienen de una mutación aleatoria durante la replicación o de una "mutación dirigida por el estrés", en la que los genes implicados en un proceso específico limitante del crecimiento tienen una mayor tasa de mutación.
Algunas bacterias también transfieren material genético entre las células. Esto puede ocurrir de tres formas principales. En primer lugar, las bacterias pueden absorber el ADN exógeno de su entorno, en un proceso llamado transformación. Muchas bacterias pueden absorber naturalmente el ADN del medio ambiente, mientras que otras deben modificarse químicamente para inducirlas a absorber el ADN. El desarrollo de la competencia en la naturaleza generalmente se asocia con condiciones ambientales estresantes, y parece ser una adaptación para facilitar la reparación del daño del ADN en las células receptoras. La segunda forma en que las bacterias transfieren material genético es mediante transducción, cuando la integración de un bacteriófago introduce ADN extraño en el cromosoma. Existen muchos tipos de bacteriófagos, algunos simplemente infectan y lisan sus bacterias hospedadoras, mientras que otros se insertan en el cromosoma bacteriano. Las bacterias resisten la infección de fagos mediante sistemas de modificación de restricción que degradan el ADN extraño y un sistema que utiliza secuencias CRISPR para retener fragmentos de los genomas de fagos con los que las bacterias han entrado en contacto en el pasado, lo que les permite bloquear la replicación del virus a través de una forma de la interferencia de ARN. El tercer método de transferencia de genes es la conjugación, mediante la cual el ADN se transfiere a través del contacto directo con las células. En circunstancias normales, la transducción, conjugación y transformación implican la transferencia de ADN entre bacterias individuales de la misma especie, pero ocasionalmente puede ocurrir transferencia entre individuos de diferentes especies bacterianas y esto puede tener consecuencias importantes, tales como la transferencia de resistencia a antibióticos. En esos casos,
Comportamiento
Movimiento

Muchas bacterias son móviles y pueden moverse usando una variedad de mecanismos. Los mejor estudiados son los flagelos, filamentos largos que un motor en la base gira para generar un movimiento parecido a la hélice. El flagelo bacteriano está compuesto por aproximadamente 20 proteínas, con aproximadamente otras 30 proteínas necesarias para su regulación y ensamblaje. El flagelo es una estructura giratoria impulsada por un motor reversible en la base que utiliza el gradiente electroquímico a través de la membrana para proporcionar potencia.

Las bacterias pueden usar flagelos de diferentes maneras para generar diferentes tipos de movimiento. Muchas bacterias (como E. coli ) tienen dos modos distintos de movimiento: avance (natación) y caída. La caída les permite reorientarse y hace que su movimiento sea una caminata aleatoria tridimensional. Las especies bacterianas difieren en el número y disposición de los flagelos en su superficie; algunos tienen un solo flagelo ( monótono ), un flagelo en cada extremo ( anfítrofo ), grupos de flagelos en los polos de la célula ( lophotrichous ), mientras que otros tienen flagelos distribuidos en toda la superficie de la célula ( peritrichous)) Los flagelos de un grupo único de bacterias, las espiroquetas, se encuentran entre dos membranas en el espacio periplásmico. Tienen un cuerpo helicoidal distintivo que se retuerce a medida que se mueve.
Otros dos tipos de movimiento bacteriano, llamados motilidad de tics y motilidad de deslizamiento, se basan en una estructura llamada pilus tipo IV. En estos tipos de motilidad, el pilus similar a una varilla se extiende desde la célula, se une a algún sustrato, y luego se retrae, tirando de la célula hacia adelante.
Las bacterias móviles son atraídas o repelidas por ciertos estímulos en comportamientos llamados impuestos : estos incluyen la quimiotaxis, la fototaxis, los taxis de energía y la magnetotaxis. En un grupo peculiar, las mixobacterias, las bacterias individuales se mueven juntas para formar ondas de células que luego se diferencian para formar cuerpos fructíferos que contienen esporas. Las mixobacterias se mueven solo cuando están sobre superficies sólidas, a diferencia de E. coli , que es móvil en medios líquidos o sólidos.
Varias especies de Listeria y Shigella se mueven dentro de las células del huésped usurpando el citoesqueleto, que normalmente se usa para mover los orgánulos dentro de la célula. Al promover la polimerización de actina en un polo de sus células, pueden formar una especie de cola que los empuja a través del citoplasma de la célula huésped.
Comunicación
Algunas bacterias tienen sistemas químicos que generan luz. Esta bioluminiscencia a menudo ocurre en bacterias que viven en asociación con peces, y la luz probablemente sirva para atraer peces u otros animales grandes.
Las bacterias a menudo funcionan como agregados multicelulares conocidos como biofilms, intercambiando una variedad de señales moleculares para la comunicación entre células, y participando en un comportamiento multicelular coordinado.
Los beneficios comunes de la cooperación multicelular incluyen una división celular del trabajo, el acceso a recursos que no pueden ser efectivamente utilizados por células individuales, la defensa colectiva contra los antagonistas y la optimización de la supervivencia de la población mediante la diferenciación en distintos tipos de células. Por ejemplo, las bacterias en las biopelículas pueden tener más de 500 veces mayor resistencia a los agentes antibacterianos que las bacterias "planctónicas" individuales de la misma especie.
Un tipo de comunicación inter-celular por una señal molecular se llama detección de quórum, que sirve para el propósito de determinar si existe una densidad de población local que es suficientemente alto que es productivo invertir en procesos que son sólo tiene éxito si un gran número de semejante los organismos se comportan de manera similar, como en excretar enzimas digestivas o emitir luz.
La detección de quórum permite a las bacterias coordinar la expresión génica y les permite producir, liberar y detectar autoinductores o feromonas que se acumulan con el crecimiento de la población celular.
Clasificación e identificación


La clasificación busca describir la diversidad de especies bacterianas al nombrar y agrupar organismos en base a similitudes. Las bacterias se pueden clasificar en función de la estructura celular, el metabolismo celular o las diferencias en los componentes celulares, como ADN, ácidos grasos, pigmentos, antígenos y quinonas. Aunque estos esquemas permitieron la identificación y clasificación de cepas bacterianas, no estaba claro si estos las diferencias representaban la variación entre especies distintas o entre cepas de la misma especie. Esta incertidumbre se debió a la falta de estructuras distintivas en la mayoría de las bacterias, así como a la transferencia lateral de genes entre especies no relacionadas. Debido a la transferencia lateral de genes, algunas bacterias estrechamente relacionadas pueden tener morfologías y metabolismos muy diferentes. Para superar esta incertidumbre, La clasificación bacteriana moderna enfatiza la sistemática molecular, utilizando técnicas genéticas como la determinación de la proporción de guanina citosina, la hibridación genoma-genoma, así como la secuenciación de genes que no se han sometido a una amplia transferencia lateral de genes, como el gen rRNA. La clasificación de las bacterias se determina mediante publicación en International Journal of Systematic Bacteriology, y Bergey's Manual of Systematic Bacteriology. El Comité Internacional de Bacteriología Sistemática (ICSB) mantiene normas internacionales para el nombramiento de bacterias y categorías taxonómicas y para su clasificación en el Código Internacional de Nomenclatura de Bacterias. como el gen rRNA. La clasificación de las bacterias se determina mediante publicación en International Journal of Systematic Bacteriology, y Bergey's Manual of Systematic Bacteriology. El Comité Internacional de Bacteriología Sistemática (ICSB) mantiene normas internacionales para el nombramiento de bacterias y categorías taxonómicas y para su clasificación en el Código Internacional de Nomenclatura de Bacterias. como el gen rRNA. La clasificación de las bacterias se determina mediante publicación en International Journal of Systematic Bacteriology, y Bergey's Manual of Systematic Bacteriology. El Comité Internacional de Bacteriología Sistemática (ICSB) mantiene normas internacionales para el nombramiento de bacterias y categorías taxonómicas y para su clasificación en el Código Internacional de Nomenclatura de Bacterias.
El término "bacteria" se aplicaba tradicionalmente a todos los procariotas microscópicos de células únicas. Sin embargo, la sistemática molecular mostró que la vida procariótica consistía en dos dominios separados, originalmente llamados Eubacteria y Archaebacteria , pero ahora se llamaba Bacteria y Archaea. que evolucionó independientemente de un ancestro común antiguo. Las arqueas y eucariotas están más estrechamente relacionadas entre sí que cualquiera de las bacterias. Estos dos dominios, junto con Eukarya, son la base del sistema de tres dominios, que actualmente es el sistema de clasificación más utilizado en microbiología. Sin embargo, debido a la introducción relativamente reciente de la sistemática molecular y un rápido aumento en el número de secuencias de genoma que están disponibles, la clasificación bacteriana sigue siendo un campo cambiante y en expansión. Por ejemplo, algunos biólogos argumentan que las Archaea y Eukaryotes evolucionaron a partir de bacterias gram-positivas.
La identificación de bacterias en el laboratorio es particularmente relevante en medicina, donde el tratamiento correcto está determinado por las especies bacterianas que causan una infección. En consecuencia, la necesidad de identificar patógenos humanos fue un gran impulso para el desarrollo de técnicas para identificar bacterias.
La tinción de Gram , desarrollada en 1884 por Hans Christian Gram, caracteriza a las bacterias en función de las características estructurales de sus paredes celulares. Las capas gruesas de peptidoglicano en la pared celular "grampositiva" se tiñen de color púrpura, mientras que la pared celular delgada "gramnegativa" aparece rosada. Combinando la morfología y la tinción de Gram, la mayoría de las bacterias pueden clasificarse como pertenecientes a uno de cuatro grupos (cocos grampositivos, bacilos grampositivos, cocos gramnegativos y bacilos gramnegativos). Algunos organismos se identifican mejor con tinciones distintas de la tinción de Gram, particularmente micobacterias o Nocardia , que muestran resistencia al ácido en Ziehl-Neelsen o manchas similares. Otros organismos pueden necesitar ser identificados por su crecimiento en medios especiales, o por otras técnicas, como la serología.
Las técnicas de cultivo están diseñadas para promover el crecimiento e identificar bacterias particulares, a la vez que restringen el crecimiento de las otras bacterias en la muestra. A menudo estas técnicas están diseñadas para especímenes específicos; por ejemplo, una muestra de esputo se tratará para identificar organismos que causan neumonía, mientras que las muestras de heces se cultivan en medios selectivos para identificar organismos que causan diarrea, al tiempo que se impide el crecimiento de bacterias no patógenas. Las muestras que normalmente son estériles, como sangre, orina o fluido espinal, se cultivan en condiciones diseñadas para desarrollar todos los organismos posibles. Una vez que se ha aislado un organismo patógeno, se puede caracterizar además por su morfología, patrones de crecimiento (como crecimiento aeróbico o anaeróbico), patrones de hemólisis y tinción.
Al igual que con la clasificación bacteriana, la identificación de bacterias está utilizando cada vez más métodos moleculares. Los diagnósticos que utilizan herramientas basadas en ADN, como la reacción en cadena de la polimerasa, son cada vez más populares debido a su especificidad y velocidad, en comparación con los métodos basados en el cultivo. Estos métodos también permiten la detección e identificación de células "viables pero no cultivables" que son metabólicamente activas pero no se dividen. Sin embargo, incluso utilizando estos métodos mejorados, se desconoce el número total de especies bacterianas y ni siquiera se pueden estimar con certeza. Siguiendo la clasificación actual, hay un poco menos de 9,300 especies conocidas de procariotas, que incluyen bacterias y arqueas;
Interacciones con otros organismos
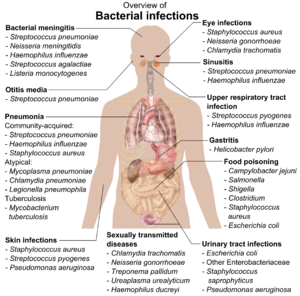
A pesar de su aparente simplicidad, las bacterias pueden formar asociaciones complejas con otros organismos. Estas asociaciones simbióticas se pueden dividir en parasitismo, mutualismo y comensalismo. Debido a su pequeño tamaño, las bacterias comensales son omnipresentes y crecen en los animales y las plantas exactamente como crecerán en cualquier otra superficie. Sin embargo, su crecimiento puede aumentar por la calidez y el sudor, y grandes poblaciones de estos organismos en los humanos son la causa del olor corporal.
Depredadores
Algunas especies de bacterias matan y luego consumen otros microorganismos, estas especies se llaman bacterias depredadoras . Estos incluyen organismos como Myxococcus xanthus , que forma enjambres de células que matan y digieren las bacterias que encuentran. Otros depredadores bacterianos se unen a su presa para digerirlos y absorber nutrientes, como Vampirovibrio chlorellavorus , o invaden otra célula y se multiplican dentro del citosol, como Daptobacter . Se cree que estas bacterias predatorias han evolucionado a partir de saprófagos que consumieron microorganismos muertos, a través de adaptaciones que les permitieron atrapar y matar a otros organismos.
Mutualistas
Ciertas bacterias forman asociaciones espaciales cercanas que son esenciales para su supervivencia. Una de estas asociaciones mutualistas, llamada transferencia interespecies de hidrógeno, se produce entre grupos de bacterias anaeróbicas que consumen ácidos orgánicos, como ácido butírico o ácido propiónico, y producen hidrógeno y arqueas metanogénicas que consumen hidrógeno. Las bacterias en esta asociación no pueden consumir los ácidos orgánicos ya que esta reacción produce hidrógeno que se acumula en su entorno. Solo la asociación íntima con Archaea, que consume hidrógeno, mantiene la concentración de hidrógeno lo suficientemente baja como para permitir que las bacterias crezcan.
En el suelo, los microorganismos que residen en la rizosfera (una zona que incluye la superficie de la raíz y el suelo que se adhiere a la raíz después de un suave movimiento) llevan a cabo la fijación de nitrógeno, convirtiendo el gas nitrógeno en compuestos nitrogenados. Esto sirve para proporcionar una forma de nitrógeno fácilmente absorbible para muchas plantas, que no pueden fijar nitrógeno por sí mismas. Muchas otras bacterias se encuentran como simbiontes en humanos y otros organismos. Por ejemplo, la presencia de más de 1.000 especies bacterianas en la flora intestinal humana normal de los intestinos puede contribuir a la inmunidad del intestino, sintetizar vitaminas, como ácido fólico, vitamina K y biotina, convertir los azúcares en ácido láctico (ver Lactobacillus), así como la fermentación de carbohidratos complejos no digeribles. La presencia de esta flora intestinal también inhibe el crecimiento de bacterias potencialmente patógenas (generalmente a través de la exclusión competitiva) y estas bacterias beneficiosas se venden como suplementos dietéticos probióticos.
Patógenos
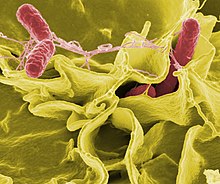
Si las bacterias forman una asociación parasitaria con otros organismos, se clasifican como patógenos. Las bacterias patógenas son una causa importante de muerte y enfermedad en los seres humanos y causan infecciones como el tétanos, la fiebre tifoidea, la difteria, la sífilis, el cólera, las enfermedades transmitidas por los alimentos, la lepra y la tuberculosis. Una causa patógena para una enfermedad médica conocida solo puede descubrirse muchos años después, como fue el caso con Helicobacter pylori y la enfermedad de úlcera péptica. Las enfermedades bacterianas también son importantes en la agricultura, con bacterias que causan manchas en las hojas, tizón de fuego y marchiteces en las plantas, así como la enfermedad de Johne, mastitis, salmonella y ántrax en los animales de granja.
Cada especie de patógeno tiene un espectro característico de interacciones con sus huéspedes humanos. Algunos organismos, como Staphylococcus o Streptococcus , pueden causar infecciones de la piel, neumonía, meningitis e incluso sepsis abrumadora, una respuesta inflamatoria sistémica que produce shock, vasodilatación masiva y muerte. Sin embargo, estos organismos también son parte de la flora humana normal y generalmente existen en la piel o en la nariz sin causar ninguna enfermedad en absoluto. Otros organismos invariablemente causan enfermedades en humanos, como Rickettsia, que son parásitos intracelulares obligados que pueden crecer y reproducirse solo dentro de las células de otros organismos. Una especie de Rickettsia causa tifus, mientras que otra causa la fiebre maculosa de las Montañas Rocosas. Chlamydia, otro filo de parásitos intracelulares obligados, contiene especies que pueden causar neumonía o infección del tracto urinario y pueden estar involucradas en la enfermedad coronaria. Finalmente, algunas especies, como Pseudomonas aeruginosa , Burkholderia cenocepacia y Mycobacterium avium , son patógenos oportunistas y causan enfermedades principalmente en personas que padecen inmunosupresión o fibrosis quística.
Las infecciones bacterianas se pueden tratar con antibióticos, que se clasifican como bactericidas si matan las bacterias, o bacteriostáticas si solo evitan el crecimiento bacteriano. Hay muchos tipos de antibióticos y cada clase inhibe un proceso que es diferente en el patógeno del que se encuentra en el huésped. Un ejemplo de cómo los antibióticos producen toxicidad selectiva son el cloranfenicol y la puromicina, que inhiben el ribosoma bacteriano, pero no el ribosoma eucariótico estructuralmente diferente. Los antibióticos se utilizan tanto en el tratamiento de enfermedades humanas como en la agricultura intensiva para promover el crecimiento de los animales, donde pueden estar contribuyendo al rápido desarrollo de la resistencia a los antibióticos en las poblaciones bacterianas. Las infecciones se pueden prevenir con medidas antisépticas, como esterilizar la piel antes de perforarla con la aguja de una jeringa, y por el cuidado apropiado de los catéteres permanentes. Los instrumentos quirúrgicos y dentales también se esterilizan para evitar la contaminación por bacterias. Los desinfectantes como el blanqueador se usan para matar bacterias u otros patógenos en las superficies para evitar la contaminación y reducir aún más el riesgo de infección.
Importancia en tecnología e industria
Las bacterias, a menudo bacterias del ácido láctico, como Lactobacillus y Lactococcus , en combinación con levaduras y mohos, se han utilizado durante miles de años en la preparación de alimentos fermentados, como queso, encurtidos, salsa de soja, chucrut, vinagre, vino y yogur.
La capacidad de las bacterias para degradar una variedad de compuestos orgánicos es notable y se ha utilizado en el procesamiento de residuos y la biorremediación. Las bacterias capaces de digerir los hidrocarburos en el petróleo a menudo se utilizan para limpiar los derrames de petróleo. Se agregó fertilizante a algunas de las playas en Prince William Sound en un intento de promover el crecimiento de estas bacterias naturales después del derrame de petróleo Exxon Valdez en 1989 . Estos esfuerzos fueron efectivos en playas que no estaban demasiado cubiertas de petróleo. Las bacterias también se usan para la biorremediación de desechos tóxicos industriales. En la industria química, las bacterias son más importantes en la producción de químicos enantioméricamente puros para su uso como productos farmacéuticos o agroquímicos.
Las bacterias también pueden usarse en lugar de pesticidas en el control biológico de plagas. Esto comúnmente implica Bacillus thuringiensis (también llamado BT), una bacteria gram-positiva que habita en el suelo. Las subespecies de esta bacteria se usan como insecticidas específicos para lepidópteros bajo nombres comerciales como Dipel y Thuricide. Debido a su especificidad, estos pesticidas se consideran inocuos para el medio ambiente, con poco o ningún efecto en los seres humanos, la vida silvestre, los polinizadores y la mayoría de los otros insectos beneficiosos.
Debido a su capacidad para crecer rápidamente y la relativa facilidad con la que pueden ser manipulados, las bacterias son el caballo de batalla para los campos de la biología molecular, la genética y la bioquímica. Al hacer mutaciones en el ADN bacteriano y examinar los fenotipos resultantes, los científicos pueden determinar la función de los genes, las enzimas y las vías metabólicas en las bacterias, y luego aplicar este conocimiento a los organismos más complejos. Este objetivo de comprender la bioquímica de una célula alcanza su expresión más compleja en la síntesis de grandes cantidades de datos enzimáticos cinéticos y de expresión génica en modelos matemáticos de organismos completos. Esto se puede lograr en algunas bacterias bien estudiadas, con modelos de Escherichia coli metabolismo ahora siendo producido y probado. Esta comprensión del metabolismo bacteriano y la genética permite el uso de la biotecnología para bioingeniería de bacterias para la producción de proteínas terapéuticas, como insulina, factores de crecimiento o anticuerpos.
Debido a su importancia para la investigación en general, las muestras de cepas bacterianas son aisladas y preservadas en los Centros de Recursos Biológicos. Esto garantiza la disponibilidad de la cepa a los científicos de todo el mundo.
Historia de la bacteriología

Las bacterias fueron observadas por primera vez por el microscopista holandés Antonie van Leeuwenhoek en 1676, utilizando un microscopio de lente única de su propio diseño. Luego publicó sus observaciones en una serie de cartas a la Royal Society of London. Las bacterias fueron el descubrimiento microscópico más notable de Leeuwenhoek. Estaban justo al límite de lo que podían distinguir sus lentes simples y, en uno de los hiatos más llamativos de la historia de la ciencia, nadie más los vería por más de un siglo. Sus observaciones también incluyeron protozoos a los que llamó animálculos, y sus hallazgos fueron examinados nuevamente a la luz de los hallazgos más recientes de la teoría celular.
Christian Gottfried Ehrenberg introdujo la palabra "bacteria" en 1828. De hecho, su Bacterium era un género que contenía bacterias en forma de vara que no formaban esporas, a diferencia de Bacillus , un género de bacterias en forma de vara que forman esporas definidas por Ehrenberg. en 1835.
Louis Pasteur demostró en 1859 que el crecimiento de microorganismos causa el proceso de fermentación, y que este crecimiento no se debe a la generación espontánea. (Las levaduras y los mohos, comúnmente asociados con la fermentación, no son bacterias, sino más bien hongos). Junto con su contemporáneo Robert Koch, Pasteur fue uno de los primeros defensores de la teoría de la enfermedad de los gérmenes.
Robert Koch, un pionero en microbiología médica, trabajó en el cólera, el ántrax y la tuberculosis. En su investigación sobre la tuberculosis, Koch finalmente demostró la teoría de los gérmenes, por la que recibió el Premio Nobel en 1905. En los postulados de Koch , estableció los criterios para probar si un organismo es la causa de una enfermedad, y estos postulados todavía se usan hoy en día.
Se dice que Ferdinand Cohn es uno de los fundadores de la bacteriología y que estudió las bacterias desde 1870. Cohn fue el primero en clasificar las bacterias en función de su morfología.
Aunque en el siglo XIX se sabía que las bacterias son la causa de muchas enfermedades, no se disponía de tratamientos antibacterianos eficaces. En 1910, Paul Ehrlich desarrolló el primer antibiótico, mediante el cambio de tintes que tiñeron selectivamente Treponema pallidum ( la espiroqueta que causa la sífilis) en compuestos que mataron selectivamente al patógeno. Ehrlich había sido galardonado con un Premio Nobel de 1908 por su trabajo en inmunología, y fue pionero en el uso de tintes para detectar e identificar bacterias, siendo su trabajo la base de la tinción de Gram y la tinción de Ziehl-Neelsen.
Un importante paso adelante en el estudio de las bacterias llegó en 1977 cuando Carl Woese reconoció que las arqueas tienen una línea separada de descendencia evolutiva de las bacterias. Esta nueva taxonomía filogenética dependió de la secuenciación del ARN ribosómico 16S y dividió los procariotas en dos dominios evolutivos, como parte del sistema de tres dominios.
Obtenido de: https://en.wikipedia.org/wiki/Bacteria