Dinosaurio
Definición
Los dinosaurios son un grupo diverso de reptiles del clado Dinosauria . Aparecieron por primera vez durante el Triassicperiod, entre 243 y 233.23 millones de años atrás, aunque el origen exacto y el momento de la evolución de los dinosaurios es objeto de una investigación activa. Se convirtieron en los vertebrados terrestres dominantes después del evento de extinción Triásico-Jurásico hace 201 millones de años; su dominio continuó a través de los períodos Jurásico y Cretácico. La ingeniería genética inversa y el registro fósil demuestran que las aves son dinosaurios emplumados modernos, que han evolucionado a partir de terópodos anteriores durante el Período Jurásico tardío. Como tal, las aves fueron el único linaje de dinosaurios que sobrevivió al evento de extinción Cretácico-Paleógeno hace 66 millones de años. Los dinosaurios se pueden dividir en dinosaurios aviares o aves; y dinosaurios no aviares , que son dinosaurios distintos de las aves. Este artículo trata principalmente con dinosaurios no aviares.Los dinosaurios son un grupo variado de animales desde puntos de vista taxonómicos, morfológicos y ecológicos. Las aves, en más de 10,000 especies vivientes, son el grupo más diverso de vertebrados además del pez perciforme. Utilizando evidencia fósil, los paleontólogos han identificado más de 500 géneros distintos y más de 1.000 especies diferentes de dinosaurios no aviares. Los dinosaurios están representados en todos los continentes por las dos especies existentes (aves) y restos fósiles. A lo largo de la primera mitad del siglo XX, antes de que las aves fueran reconocidas como dinosaurios, la mayoría de la comunidad científica creía que los dinosaurios eran lentos y de sangre fría. La mayoría de las investigaciones realizadas desde la década de 1970, sin embargo, han indicado que todos los dinosaurios eran animales activos con metabolismos elevados y numerosas adaptaciones para la interacción social. Algunos eran herbívoros, otros carnívoros.

Una colección de esqueletos de dinosaurios fósiles. En el sentido de las agujas del reloj desde arriba a la izquierda: Microraptor gui (un terópodo alado), Apatosaurus louisae (un saurópodo gigante), Edmontosaurus regalis (un ornitópodo pico de pato), Triceratops horridus (un ceratopsio cornudo), Stegosaurus stenops (un estegaurio chapado), Pinacosaurus grangeri ( un anquilosaurio blindado)
Una colección de esqueletos de dinosaurios fósiles. En el sentido de las agujas del reloj desde arriba a la izquierda: Microraptor gui (un terópodo alado), Apatosaurus louisae (un saurópodo gigante), Edmontosaurus regalis (un ornitópodo pico de pato), Triceratops horridus (un ceratopsio cornudo), Stegosaurus stenops (un estegaurio chapado), Pinacosaurus grangeri ( un anquilosaurio blindado)
Mientras que los dinosaurios eran ancestralmente bípedos, muchos grupos extintos incluyeron especies cuadrúpedas, y algunos fueron capaces de cambiar entre estas posturas. Las estructuras de visualización elaboradas, como los cuernos o las crestas, son comunes a todos los grupos de dinosaurios, y algunos grupos extintos desarrollaron modificaciones esqueléticas, como armaduras óseas y espinas. Mientras que el linaje aviar de los dinosaurios (aves) en la actualidad es generalmente pequeño debido a las limitaciones del vuelo, muchos dinosaurios prehistóricos (no aviarios y aviares) tenían cuerpo grande: se estima que los dinosaurios saurópodos más grandes alcanzaron longitudes de 39,7. metros (130 pies) y alturas de 18 metros (59 pies) y fueron los animales terrestres más grandes de todos los tiempos. Aún así, la idea de que los dinosaurios no aviares eran uniformemente gigantescos es una idea errónea basada en parte en el sesgo de preservación, tan grande, los huesos fuertes son más propensos a durar hasta que se fosilicen. Muchos dinosaurios eran bastante pequeños: Xixianykus , por ejemplo, tenía solo unos 50 cm (20 in) de largo.
Desde que se reconocieron los primeros fósiles de dinosaurios a principios del siglo XIX, los esqueletos montados de dinosaurios fósiles han sido importantes atracciones en museos de todo el mundo, y los dinosaurios se han convertido en una parte perdurable de la cultura mundial. Los grandes tamaños de algunos grupos de dinosaurios, así como su naturaleza aparentemente monstruosa y fantástica, han asegurado la aparición regular de los dinosaurios en los libros y películas más vendidos, como Jurassic Park . El persistente entusiasmo del público por los animales ha resultado en fondos significativos para la ciencia de los dinosaurios, y los nuevos descubrimientos son cubiertos regularmente por los medios.
Etimología
El taxón Dinosauria fue formalmente nombrado en 1841 por el paleontólogo Sir Richard Owen, quien lo usó para referirse a la "tribu distinta o suborden de los reptiles saurios" que luego se reconocía en Inglaterra y en todo el mundo. El término se deriva del griego antiguo δεινός (deinos) , que significa "terrible, potente o terriblemente grande", y σαῦρος (sauros) , que significa 'lagarto o reptil'. Aunque el nombre taxonómico a menudo se ha interpretado como una referencia a los dientes de los dinosaurios, garras y otras características temibles, Owen pretendió simplemente para evocar su tamaño y majestuosidad.
Otros animales prehistóricos, incluidos mosasaurios, ictiosaurios, pterosaurios, plesiosaurios y Dimetrodon , aunque a menudo se los concibe popularmente como dinosaurios, no se clasifican taxonómicamente como dinosaurios.
Definición

Bajo la nomenclatura filogenética, los dinosaurios se definen generalmente como el grupo que consiste en el ancestro común más reciente (MRCA) de Triceratops y Neornithes, y todos sus descendientes. También se ha sugerido que Dinosauria se defina con respecto al MRCA de Megalosaurus e Iguanodon, porque estos fueron dos de los tres géneros citados por Richard Owen cuando reconoció a los Dinosauria. Ambas definiciones dan como resultado el mismo conjunto de animales definidos como dinosaurios: "Dinosauria = Ornithischia + Saurischia", que abarca anquilosaurios (cuadrúpedos herbívoros acorazados), estegosaurios (cuadrúpedos herbívoros plateados), ceratópsidos (cuadrúpedos herbívoros con cuernos y adornos), ornitópodos (bípedos) o herbívoros cuadrúpedos que incluyen "pato-billetes"), terópodos (en su mayoría bípedos carnívoros y aves) y sauropodomorfos (en su mayoría grandes cuadrúpedos herbívoros con cuellos y colas largas).
Ahora se reconoce que las aves son el único linaje sobreviviente de los dinosaurios terópodos. En la taxonomía tradicional, las aves se consideraban una clase separada que había evolucionado de los dinosaurios, un superorden distinto. Sin embargo, la mayoría de los paleontólogos contemporáneos preocupados por los dinosaurios rechazan el estilo tradicional de clasificación a favor de la taxonomía filogenética; este enfoque requiere que, para que un grupo sea natural, todos los descendientes de los miembros del grupo también deben ser incluidos en el grupo. Por lo tanto, se considera que los pájaros son dinosaurios y, por lo tanto, los dinosaurios no están extintos. Las aves se clasifican como pertenecientes al subgrupo Maniraptora, que son celurosaurios, que son terópodos, que son saurisquios, que son dinosaurios.
La investigación de Matthew Baron, David B. Norman y Paul M. Barrett en 2017 sugirió una revisión radical de la sistemática de dinosaurios. Análisis filogenético por Baron et al. Recuperó Ornithischia como más cercano a Theropoda que Sauropodomorpha, en comparación con la unión tradicional de terópodos con sauropodomorphs. Resucitaron el clado Ornithoscelida para referirse al grupo que contiene Ornithischia y Theropoda. La propia dinosauria fue redefinida como el último ancestro común de Triceratops horridus , Passer domesticus , Diplodocus carnegii y todos sus descendientes, para asegurar que los saurópodos y los parientes permanezcan incluidos como dinosaurios.
Descripción general

Utilizando una de las definiciones anteriores, los dinosaurios se pueden describir generalmente como arcosaurios con extremidades posteriores mantenidas erectas debajo del cuerpo. Muchos grupos de animales prehistóricos se conciben popularmente como dinosaurios, como ictiosauros, mosasaurios, plesiosaurios, pterosaurios y pelicosaurios (especialmente Dimetrodon ), pero no se clasifican científicamente como dinosaurios, y ninguno tenía la postura erguida de la extremidad posterior característica de los verdaderos dinosaurios. Los dinosaurios fueron los vertebrados terrestres dominantes del Mesozoico, especialmente los períodos Jurásico y Cretácico. Otros grupos de animales fueron restringidos en tamaño y nichos; los mamíferos, por ejemplo, raramente excedían el tamaño de un gato doméstico, y generalmente eran carnívoros del tamaño de un roedor de presas pequeñas.
Los dinosaurios siempre han sido un grupo extremadamente variado de animales; según un estudio de 2006, más de 500 géneros de dinosaurios no aviares se han identificado con certeza hasta ahora, y el número total de géneros conservados en el registro fósil se ha estimado en alrededor de 1850, de los cuales casi el 75% aún no se han descubierto. Un estudio anterior predijo que existían unos 3.400 géneros de dinosaurios, incluidos muchos que no se habrían conservado en el registro fósil. Para el 17 de septiembre de 2008, se habían nombrado 1.047 especies diferentes de dinosaurios.
En 2016, el número estimado de especies de dinosaurios que existían en la era Mesozoica se estimó en 1,543-2,468. Algunos son herbívoros, otros carnívoros, incluidos los que comen semillas, los que comen peces, los insectívoros y los omnívoros. Mientras que los dinosaurios eran ancestralmente bípedos (como lo son todas las aves modernas), algunas especies prehistóricas eran cuadrúpedos, y otros, como Anchisaurus e Iguanodon, podría caminar con la misma facilidad en dos o cuatro patas. Las modificaciones craneales como cuernos y crestas son rasgos dinosaurios comunes, y algunas especies extintas tenían una armadura ósea. Aunque conocidos por su gran tamaño, muchos dinosaurios mesozoicos eran de tamaño humano o más pequeños, y las aves modernas generalmente son de tamaño pequeño. Los dinosaurios de hoy habitan en todos los continentes, y los fósiles muestran que habían logrado una distribución global al menos al principio del período Jurásico. Las aves modernas habitan en la mayoría de los hábitats disponibles, desde terrestres hasta marinos, y hay evidencia de que algunos dinosaurios no aviares (como Microraptor ) podían volar o al menos deslizarse, y otros, como los espinosáuridos, tenían hábitos semiacuáticos.
Distinguir las características anatómicas
Mientras que los descubrimientos recientes han hecho más difícil presentar una lista universalmente acordada de las características distintivas de los dinosaurios, casi todos los dinosaurios descubiertos hasta ahora comparten ciertas modificaciones al esqueleto arosaurios ancestral, o son claros descendientes de dinosaurios más antiguos que muestran estas modificaciones. Aunque algunos grupos posteriores de dinosaurios presentaron versiones modificadas de estos rasgos, se consideran típicos de Dinosauria; los primeros dinosaurios los tenían y se los pasaron a sus descendientes. Tales modificaciones, que se originan en el ancestro común más reciente de cierto grupo taxonómico, se llaman sinapomorfias de dicho grupo.
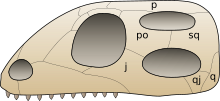
Una evaluación detallada de las interrelaciones de los arcosáuridos por Sterling Nesbitt confirmó o encontró las siguientes doce sinapomorfías no ambiguas, algunas previamente conocidas:
- en el cráneo, una fosa supratemporal (excavación) está presente frente a la fenestra supratemporal, la abertura principal en el techo del cráneo posterior
- epífisis, procesos de apunte oblicuo hacia atrás en las esquinas superiores posteriores, presentes en las vértebras cervicales anteriores (delanteras) detrás del atlas y el eje, las dos primeras vértebras del cuello
- ápice de la cresta deltopectoral (una proyección en la que se unen los músculos deltopectoral) ubicada en o más del 30% a lo largo de la longitud del húmero (hueso del brazo superior)
- radio, un hueso del brazo inferior, más corto que el 80% de la longitud del húmero
- cuarto trocánter (proyección donde el músculo caudofemoralis se une en el eje trasero interno) en el fémur (hueso del muslo) es una pestaña filosa
- cuarto trocánter asimétrico, con margen distal inferior que forma un ángulo más pronunciado con respecto al eje
- en el astrágalo y el calcáneo, los huesos del tobillo superior, la faceta articular proximal, la superficie de conexión superior, para el peroné ocupa menos del 30% de la anchura transversal del elemento
- los exoccipitales (huesos en la parte posterior del cráneo) no se encuentran a lo largo de la línea media en el piso de la cavidad endocraneal, el espacio interno de la caja craneal
- en la pelvis, las superficies articulares proximales del isquion con el ilion y el pubis están separadas por una gran superficie cóncava (en la parte superior del isquion una parte de la articulación de la cadera abierta se encuentra entre los contactos con el hueso púbico y el ilion)
- cresta cnemial en la tibia (parte sobresaliente de la superficie superior de la tibia) arcos anterolateralmente (curvas hacia el frente y el lado exterior)
- cresta distinta orientada proximalmente (vertical) presente en la cara posterior del extremo distal de la tibia (la superficie posterior del extremo inferior de la tibia)
- superficie articular cóncava para el peroné del calcáneo (la superficie superior del calcáneo, donde toca el peroné, tiene un perfil hueco)
Nesbitt encontró varias sinapomorfias potenciales adicionales, y descontó una cantidad de sinapomorfías sugeridas anteriormente. Algunos de estos también están presentes en silesaurids, que Nesbitt recuperó como un grupo hermano de Dinosauria, incluyendo un trocánter anterior grande, metatarsianos II y IV de longitud subigual, contacto reducido entre isquion y pubis, la presencia de una cresta cnemial en la tibia y de un proceso ascendente en el astrágalo, y muchos otros.
Los dinosaurios comparten una variedad de otras características esqueléticas. Sin embargo, debido a que son comunes a otros grupos de arcosaurios o no estuvieron presentes en todos los dinosaurios tempranos, estas características no se consideran sinapomorfias. Por ejemplo, como diapsidos, los dinosaurios ancestralmente tenían dos pares de fenestras temporales (aberturas en el cráneo detrás de los ojos), y como miembros del grupo de diápsidos Archosauria, tenían aberturas adicionales en el hocico y la mandíbula inferior. Además, ahora se sabe que varias características que antes se consideraban sinapomorfías aparecieron antes que los dinosaurios, o que estuvieron ausentes en los primeros dinosaurios y evolucionaron independientemente por diferentes grupos de dinosaurios. Estos incluyen una escápula alargada, o omóplato; un sacro compuesto por tres o más vértebras fusionadas (tres se encuentran en algunos otros arcosaurios, pero solo dos se encuentran en Herrerasaurus ); y un acetábulo perforado, o cavidad de la cadera, con un orificio en el centro de su superficie interna (cerrado en Saturnalia , por ejemplo). Otra dificultad para determinar características distintivamente dinosaurios es que los dinosaurios primitivos y otros archosaurios del Triásico tardío a menudo son poco conocidos y fueron similares en muchos sentidos; estos animales algunas veces han sido mal identificados en la literatura.

Los dinosaurios están de pie con sus extremidades traseras erectas de una manera similar a la mayoría de los mamíferos modernos, pero distinta de la mayoría de los otros reptiles, cuyas extremidades se extienden a ambos lados. Esta postura se debe al desarrollo de un receso orientado lateralmente en la pelvis (generalmente un receptáculo abierto) y una cabeza distinta que se dirige hacia dentro en el fémur. Su postura erecta permitió a los dinosaurios tempranos respirar fácilmente mientras se movían, lo que probablemente permitió resistencia y niveles de actividad que superaron a los de los reptiles "en expansión". Las extremidades erectas probablemente también ayudaron a soportar la evolución de un gran tamaño al reducir las tensiones de flexión en las extremidades. Algunos archosaurios no dinosaurios, incluidos los rauisuquios, también tenían extremidades erectas, pero lograron esto mediante una configuración de "pilar erecto" de la articulación de la cadera,
Historia Evolutiva
Orígenes y evolución temprana
Los dinosaurios divergieron de sus antepasados arcosaurios durante el período triásico medio o tardío, aproximadamente 20 millones de años después de que el evento de extinción del Pérmico-Triásico aniquilara aproximadamente el 95% de toda la vida en la Tierra. La datación radiométrica de la formación rocosa que contenía fósiles del género de los primeros dinosaurios Eoraptor, con 231,4 millones de años de antigüedad, establece su presencia en el registro fósil en este momento. Los paleontólogos creen que Eoraptor se asemeja al ancestro común de todos los dinosaurios; si esto es cierto, sus rasgos sugieren que los primeros dinosaurios fueron pequeños depredadores bípedos. El descubrimiento de ornithodirans primitivos, como dinosaurios, como Marasuchus y Lagerpeton en los estratos del Triásico Medio Argentino apoya esta visión; el análisis de los fósiles recuperados sugiere que estos animales eran de hecho pequeños depredadores bípedos. Los dinosaurios pueden haber aparecido ya hace 243 millones de años, como lo demuestran los restos del género Nyasasaurus de ese período, aunque los fósiles conocidos de estos animales son demasiado fragmentarios como para saber si son dinosaurios o parientes dinosaurios muy cercanos. Recientemente, se ha determinado que el Staurikosaurus de la Formación Santa Maria data de 233.23 Ma, haciéndolo más viejo en edad geológica que Eoraptor .
Cuando aparecieron los dinosaurios, no eran los animales terrestres dominantes. Los hábitats terrestres fueron ocupados por varios tipos de archosauromorfos y terápsidos, como cinodontes y rinososaurios. Sus principales competidores fueron los pseudosuchia, como los aetosaurios, los ornitosúquidos y los rauisuquios, que tuvieron más éxito que los dinosaurios. La mayoría de estos otros animales se extinguieron en el Triásico, en uno de dos eventos. En primer lugar, hace unos 215 millones de años, una variedad de archosauromorfos basales, incluidos los protorosaurios, se extinguieron. Esto fue seguido por el evento de extinción Triásico-Jurásico (hace unos 200 millones de años), que vio el final de la mayoría de los otros grupos de los primeros arcosaurios, como los aetosaurios, los ornitosúpsidos, los fitosaurios y los rauisuquios. Rhynchosaurs y dicinodontos sobrevivieron (al menos en algunas áreas) por lo menos hasta principios de mediados de Norian y principios de Rhaetian, respectivamente, y la fecha exacta de su extinción es incierta. Estas pérdidas dejaron una fauna terrestre de crocodylomorphs, dinosaurios, mamíferos, pterosaurios y tortugas. Las primeras líneas de los primeros dinosaurios se diversificaron a través de los estadios carniense y noriense del Triásico, posiblemente al ocupar los nichos de los grupos que se extinguieron. También, en particular, hubo una mayor tasa de extinción durante el Evento Pluvial de Carnian. posiblemente ocupando los nichos de los grupos que se extinguieron. También, en particular, hubo una mayor tasa de extinción durante el Evento Pluvial de Carnian. posiblemente ocupando los nichos de los grupos que se extinguieron. También, en particular, hubo una mayor tasa de extinción durante el Evento Pluvial de Carnian.
Evolución y paleobiogeografía
La evolución de los dinosaurios después del Triásico sigue los cambios en la vegetación y la ubicación de los continentes. A fines del Triásico y principios del Jurásico, los continentes estaban conectados como la única masa de tierra Pangea, y había una fauna mundial de dinosaurios compuesta en su mayoría de carnívoros coelofídicos y herbívoros sauropodomorfos tempranos. Las plantas de Gimnosperma (especialmente coníferas), una fuente potencial de alimento, irradiaban Triásico tardío. Los primeros sauropodomorfos no tenían mecanismos sofisticados para procesar los alimentos en la boca, por lo que deben haber empleado otros medios para descomponer los alimentos a lo largo del tracto digestivo. La homogeneidad general de las faunas dinosaurianas continuó en el Jurásico medio y tardío, donde la mayoría de las localidades tenían depredadores compuestos por ceratosaurios, spinosauroides y carnosaurios, y herbívoros que consisten en ornitisquios estegosaurios y grandes saurópodos. Ejemplos de esto incluyen la Formación Morrison de América del Norte y las Camas Tendaguru de Tanzania. Los dinosaurios en China muestran algunas diferencias, con terópodos sinraptóridos especializados y saurópodos inusuales de cuello largo, como Mamenchisaurus. Los anquilosaurios y los ornitópodos también se estaban volviendo más comunes, pero los prosaurópodos se habían extinguido. Las coniferas y pteridofitas fueron las plantas más comunes. Los saurópodos, al igual que los prosaurópodos anteriores, no eran procesadores orales, pero los ornitisquios desarrollaban diversos medios para tratar los alimentos en la boca, incluidos posibles órganos parecidos a las mejillas para mantener los alimentos en la boca y movimientos de la mandíbula para moler los alimentos. Otro evento evolutivo notable del Jurásico fue la aparición de pájaros verdaderos, descendientes de los manuropácidos coelurosaurios.

A principios del Cretácico y la ruptura en curso de Pangea, los dinosaurios se diferenciaban fuertemente por la masa de tierra. La primera parte de este tiempo vio la expansión de anquilosaurios, iguanodontes y braquiosáuridos a través de Europa, América del Norte y el norte de África. Estos fueron luego complementados o reemplazados en África por grandes spinosaurid y carcharodontosauridtheropods, y rebachisaurid y titanosaurios saurópodos, también se encuentran en América del Sur. En Asia, los manuropácidos coelurosaurios como los dromeosáuridos, los troodóntidos y los oviraptoresáuridos se convirtieron en los terópodos comunes, y los anquilosáuridos y los primeros ceratópsidos como Psittacosaurio se convirtieron en herbívoros importantes Mientras tanto, Australia albergaba una fauna de anquilosaurios basales, hipsilofodontes e iguanodontes. Los estegosaurios parecen haberse extinguido en algún momento a fines del Cretácico temprano o al principio del Cretácico tardío. Un cambio importante en el Cretácico temprano, que se amplificaría a finales del Cretácico, fue la evolución de las plantas con flores. Al mismo tiempo, varios grupos de herbívoros dinosaurios desarrollaron formas más sofisticadas de procesar alimentos por vía oral. Los ceratópsidos desarrollaron un método de corte con dientes apilados el uno del otro en baterías, y los iguanodontes refinaron un método de molienda con baterías dentales, llevado a su extremo en hadrosauridos. Algunos saurópodos también desarrollaron baterías dentales, mejor ejemplificadas por el Nigersaurus rebbachisaurid .

Había tres faunas de dinosaurios generales en el Cretácico superior. En los continentes septentrionales de América del Norte y Asia, los principales terópodos eran tiranosaurios y varios tipos de terópodos maniraptoranos más pequeños, con un conjunto herbívoro predominantemente ornitisquio de hadrosáuridos, ceratópsidos, anquilosáuridos y paquicefalosaurios. En los continentes del sur, que formaban el ahora dividido Gondwana, los abelisáuridos eran los terópodos comunes, y los saurópodos titanosaurios los herbívoros comunes. Finalmente, en Europa, los dromeosáuridos, los iguanodontes rabdodóntidos, los anquilosaurios nodosauridos y los saurópodos titanosaurios prevalecieron. Las plantas con flores irradiaban mucho, y las primeras hierbas aparecían a fines del Cretáceo. Los hadrosáuridos de trituración y los ceratópsidos cortantes se volvieron extremadamente diversos en América del Norte y Asia.
El evento de extinción Cretácico-Paleógeno, que ocurrió hace aproximadamente 66 millones de años al final del período Cretácico, causó la extinción de todos los grupos de dinosaurios a excepción de las aves neornitinas. Algunos otros grupos de diápsidos, como cocodrílidos, sebecosuquios, tortugas, lagartos, serpientes, esfenodontes y coristoderanos, también sobrevivieron al evento.
Los linajes supervivientes de aves neornitinas, incluidos los ancestros de ratites, patos y pollos modernos, y una variedad de aves acuáticas, se diversificaron rápidamente al comienzo del período Paleógeno, entrando en nichos ecológicos que quedaron vacantes por la extinción de grupos de dinosaurios mesozoicos como el arbóreo enantiornitinas, hesperornitinas acuáticas e incluso los terópodos terrestres más grandes (en forma de Gastornis), eogruiids, bathornithids, ratites, geranoidids, mihirungs, y "terror birds"). A menudo se cita que los mamíferos compitieron con las neomitinas por el dominio de la mayoría de los nichos terrestres, pero muchos de estos grupos coexistieron con faunas ricas en mamíferos durante la mayor parte del Cenozoico. Las aves terrenales y los batorítidos ocuparon gremios carnívoros junto a mamíferos predadores, y las corredoras aún tienen bastante éxito como herbívoros medianos; Las eogruidas duraron de manera similar desde el Eoceno hasta el Plioceno, y se extinguieron muy recientemente después de más de 20 millones de años de coexistencia con muchos grupos de mamíferos.
Clasificación
Los dinosaurios pertenecen a un grupo conocido como archosaurios, que también incluye cocodrilos modernos. Dentro del grupo de los arcosaurios, los dinosaurios se diferencian más notablemente por su forma de andar. Las patas de dinosaurio se extienden directamente debajo del cuerpo, mientras que las patas de los lagartos y los cocodrílidos se extienden a ambos lados.
En conjunto, los dinosaurios como un clado se dividen en dos ramas principales, Saurischia y Ornithischia. Saurischia incluye aquellos taxones que comparten un ancestro común más reciente con aves que con Ornithischia, mientras que Ornithischia incluye todos los taxones que comparten un ancestro común más reciente con Triceratops que con Saurischia. Anatómicamente, estos dos grupos se pueden distinguir más notablemente por su estructura pélvica. Early saurischians- "-lagarto cuatro aguas", del griego sauros (σαῦρος) que significa "lagarto" y ischion (ἰσχίον) significa "articulación de la cadera" -retained la estructura de la cadera de sus antepasados, con un hueso pubis dirigida cranealmente, o hacia adelante., therizinosauroids, dromaeosaurids, y pájaros). Saurischia incluye los terópodos (exclusivamente bípedos y con una amplia variedad de dietas) y sauropodomorfos (herbívoros de cuello largo que incluyen grupos avanzados cuadrúpedos).
Por el contrario, ornithischians- "pájaro de caderas", del griego ornitheios (ὀρνίθειος) que significa "de un pájaro" y ischion (ἰσχίον) que significa "articulación de la cadera" -Tuvimos una pelvis que se asemejaba superficialmente la pelvis de un pájaro: se orientó el hueso púbico caudalmente (apuntando hacia atrás). A diferencia de las aves, el pubis ornitisquio también usualmente tenía un proceso adicional orientado hacia adelante. Ornithischia incluye una variedad de especies que fueron principalmente herbívoros. ( NB: los términos "cadera de lagarto" y "cadera de pájaro" son nombres equivocados, aves evolucionadas de dinosaurios con "caderas de lagarto").
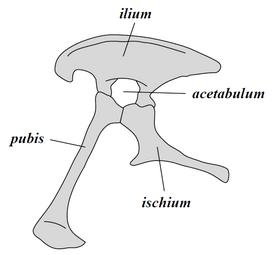 Estructura de la pelvis de Saurischian (lado izquierdo)
Estructura de la pelvis de Saurischian (lado izquierdo) Tiranosaurio pelvis (que muestra la estructura saurisquiana - lado izquierdo)
Tiranosaurio pelvis (que muestra la estructura saurisquiana - lado izquierdo) Estructura de la pelvis de Ornithischian (lado izquierdo)
Estructura de la pelvis de Ornithischian (lado izquierdo) Pelvis de Edmontosaurus (que muestra la estructura ornitisquiana - lado izquierdo)
Pelvis de Edmontosaurus (que muestra la estructura ornitisquiana - lado izquierdo)
Taxonomía
La siguiente es una clasificación simplificada de grupos de dinosaurios basada en sus relaciones evolutivas, y organizada en base a la lista de especies de dinosaurios mesozoicos proporcionadas por Holtz (2007). Una versión más detallada se puede encontrar en la clasificación de dinosaurios. La daga (†) se usa para significar grupos sin miembros vivos.
- Dinosauria
- Saurischia ("lagarto-hipped"; incluye Theropoda y Sauropodomorpha)
- Theropoda (todos bípedos, la mayoría eran carnívoros)

-
-
- † Herrerasauria (primeros carnívoros bípedos)
- † Coelophysoidea (pequeños terópodos tempranos, incluye Coelophysis y parientes cercanos)
- † Dilophosauridae (terópodos con cresta temprana y carnívoros)
- † Ceratosauria (generalmente con cuernos elaborados, los carnívoros sureños dominantes del Cretácico)
- Tetanurae ("colas rígidas"; incluye a la mayoría de los terópodos)
- † Megalosauroidea (grupo inicial de grandes carnívoros, incluidos los espinosáuridos semiacuáticos)
- † Carnosauria ( Allosaurus y parientes cercanos, como Carcharodontosaurus )
- Coelurosauria (terópodos emplumados, con una variedad de tamaños y nichos corporales)
- † Compsognathidae (primeros coelurosaurios con extremidades anteriores reducidas)
- † Tyrannosauridae ( Tyrannosaurus y parientes cercanos, con extremidades anteriores reducidas)
- † Ornithomimosauria ("avestruz-imita", en su mayoría sin dientes, carnívoros a posibles herbívoros)
- † Alvarezsauroidea (pequeños insectívoros con extremidades anteriores reducidas, cada uno con una garra agrandada)
- Maniraptora ("secuestradores de manos"; tenía brazos y dedos largos y delgados)
- † Therizinosauria (herbívoros bípedos con grandes garras y pequeñas cabezas)
- † Oviraptorosauria (en su mayoría sin dientes, su dieta y estilo de vida son inciertos)
- † Archaeopterygidae (pequeños terópodos alados o pájaros primitivos)
- † Deinonychosauria (de tamaño pequeño a mediano, parecido a un pájaro, con una garra distintiva del dedo del pie)
- Avialae (pájaros modernos y parientes extintos)
- † Scansoriopterygidae (pequeños avialanos primitivos con tres dedos largos)
- † Omnivoropterygidae (avialans grandes de cola corta temprana)
- † Confuciusornithidae (pequeños avialanos sin dientes)
- † Enantiornithes (avialanos voladores primitivos que viven en los árboles)
- Euornithes (aves voladoras avanzadas)
- † Yanornithiformes (pájaros chinos cretáceos dentados)
- † Hesperornithes (aves especializadas en buceo acuático)
- Aves (pájaros modernos con pico y sus parientes extintos)
-

-
- † Sauropodomorpha (herbívoros con cabezas pequeñas, cuellos largos, colas largas)
- † Guaibasauridae (sauropodomorfos pequeños, primitivos, omnívoros)
- † Plateosauridae (primitivos, estrictamente bípedos "prosaurópodos")
- † Riojasauridae (sauropodomorfos pequeños y primitivos)
- † Massospondylidae (sauropodomorfos pequeños y primitivos)
- † Sauropoda (muy grande y pesada, generalmente más de 15 m (49 pies) de largo, cuadrúpedo)
- † Vulcanodontidae (saurópodos primitivos con extremidades pilares)
- † Eusauropoda ("saurópodos verdaderos")
- † Cetiosauridae ("reptiles de ballena")
- † Turiasauria (grupo europeo de saurópodos del Jurásico y Cretáceo)
- † Neosauropoda ("nuevos saurópodos")
- † Diplodocoidea (calaveras y colas alargadas, dientes típicamente angostos y semejantes a un lápiz)
- † Macronaria (cráneos cuadrados, dientes con forma de cuchara o de lápiz)
- † Brachiosauridae (macronarios de largo cuello y brazos largos)
- † Titanosauria (diversa, robusta, con caderas anchas, más común en el Cretácico superior de los continentes del sur)

- † Ornithischia ("pájaro-hipped"; diversos herbívoros bípedos y cuadrúpedos)
- † Heterodontosauridae (pequeños herbívoros / omnívoros ornitópodos basales con dientes prominentes tipo canino)
- † Thyreophora (dinosaurios acorazados, en su mayoría cuadrúpedos)
- † Anquilosaurios (escudos como armadura primaria, algunos tenían colas en forma de maza)
- † Stegosauria (picos y placas como armadura primaria)
- † Neornithischia ("nuevos ornitisquios")
- † Ornithopoda (varios tamaños, bípedos y cuadrúpedos, desarrolló un método de masticar usando flexibilidad de cráneo y numerosos dientes)
- † Marginocephalia (caracterizada por un crecimiento craneal)
- † Pachycephalosauria (bípedos con crecimiento abovedado o nudoso en cráneos)
- † Ceratopsia (cuadrúpedos con volantes, muchos también tenían cuernos)
Biología
El conocimiento sobre los dinosaurios se deriva de una variedad de registros fósiles y no fósiles, que incluyen huesos fosilizados, heces, rastros, gastrolitos, plumas, impresiones de la piel, órganos internos y tejidos blandos. Muchos campos de estudio contribuyen a nuestra comprensión de los dinosaurios, incluida la física (especialmente la biomecánica), la química, la biología y las ciencias de la tierra (de las cuales la paleontología es una subdisciplina). Dos temas de particular interés y estudio han sido el tamaño y el comportamiento de los dinosaurios.
tamaño

La evidencia actual sugiere que el tamaño promedio de los dinosaurios varió a través del Triásico, el Jurásico temprano, el Jurásico tardío y el Cretácico. Dinosaurios terópodos depredadores, que ocupó la mayoría de los nichos carnívoro terrestre durante el Mesozoico, más a menudo caen en los 100 a 1 000 kg (220 a 2 200 lb) categoría cuando ordenados por peso estimado en categorías basadas en orden de magnitud, mientras que los últimos mamíferos carnivoran depredadores pico en la categoría de 10 a 100 kg (22 a 220 lb). El modo de las masas corporales del dinosaurio mesozoico es de entre una y diez toneladas métricas. Esto contrasta fuertemente con el tamaño de los mamíferos Cenozoicos, estimado por el Museo Nacional de Historia Natural en aproximadamente 2 a 5 kg (4.4 a 11.0 lb).
Los saurópodos eran los dinosaurios más grandes y pesados. Durante gran parte de la era de los dinosaurios, los saurópodos más pequeños eran más grandes que cualquier otra cosa en su hábitat, y los más grandes eran una orden de magnitud más masiva que cualquier otra cosa que haya recorrido la Tierra desde entonces. Mamíferos prehistóricos gigantes como Paraceratherium (los mamíferos terrestres más grandes) fueron eclipsados por los saurópodos gigantes, y solo las ballenas modernas se acercan o les sobrepasan en tamaño. Existen varias ventajas propuestas para el gran tamaño de los saurópodos, incluida la protección contra la depredación, la reducción del uso de energía y la longevidad, pero puede ser que la ventaja más importante sea la dieta. Los animales grandes son más eficientes en la digestión que los animales pequeños, porque los alimentos pasan más tiempo en sus sistemas digestivos. Esto también les permite subsistir con alimentos con un menor valor nutritivo que los animales más pequeños. Los restos de saurópodos se encuentran principalmente en formaciones rocosas que se interpretan como secas o estacionalmente secas, y la capacidad de comer grandes cantidades de barridos con pocos nutrientes habría sido ventajosa en dichos entornos.
Más grande y más pequeño
Los científicos probablemente nunca estarán seguros de que los dinosaurios más grandes y más pequeños jamás hayan existido. Esto se debe a que solo un pequeño porcentaje de animales se fosilizan y la mayoría de ellos permanecen enterrados en la tierra. Pocos de los especímenes que se recuperan son esqueletos completos, y las impresiones de la piel y otros tejidos blandos son raras. Reconstruir un esqueleto completo comparando el tamaño y la morfología de los huesos con los de especies similares y más conocidas es un arte inexacto, y reconstruir los músculos y otros órganos del animal vivo es, en el mejor de los casos, un proceso de conjeturas educadas.

El dinosaurio más alto y más pesado conocido por los buenos esqueletos es el brancai jirafa (anteriormente clasificado como una especie de Brachiosaurus ). Sus restos fueron descubiertos en Tanzania entre 1907 y 1912. Los huesos de varias personas de tamaño similar se incorporaron al esqueleto ahora montado y en exhibición en el Museum für Naturkunde Berlin; este monte tiene 12 metros (39 pies) de alto y 21.8-22.5 metros (72-74 pies) de largo, y habría pertenecido a un animal que pesaba entre 30 000 y 60 000 kilogramos ( 70 000 y 130 000 lb). El dinosaurio completo más largo es el Diplodocus de 27 metros (89 pies) de largo , que fue descubierto en Wyoming en los Estados Unidos y exhibido en el Carnegie Natural History Museum de Pittsburgh en 1907. El dinosaurio más largo conocido de un buen material fósil es el Patagotitan : el esqueleto en el Museo Americano de Historia Natural es de 37 metros (121 pies) largo. El Museo Carmen Funes tiene un esqueleto reconstruido Argentinosaurus de 39.7 metros (130 pies) de largo.

Había dinosaurios más grandes, pero el conocimiento de ellos se basa completamente en una pequeña cantidad de fósiles fragmentarios. La mayoría de los especímenes herbívoros más grandes registrados se descubrieron en la década de 1970 o más tarde, e incluyen el Argentinosaurus masivo , que pudo haber pesado entre 80 000 y 100 000 kilogramos (90 a 110 toneladas cortas) y alcanzó una longitud de 30-40 metros (98- 131 pies); algunos de los más largos fueron los 33.5 metros (110 pies) de largo Diplodocus hallorum (anteriormente Seismosaurus ), los 33-34 metros (108-112 pies) de largo Supersaurus y 37 metros (121 pies) de largo Patagotitan ; y el Sauroposeidon más alto, de 18 metros (59 pies) de altura , que podría haber llegado a una ventana del sexto piso. El dinosaurio más pesado y más largo puede haber sido Amphicoelias fragillimus , conocido solo por un arco neural vertebral parcial ahora perdido descrito en 1878. Extrapolando de la ilustración de este hueso, el animal pudo haber tenido 58 metros (190 pies) de largo y pesado 122 400 kg ( 270 000 lb). Sin embargo, como no se han encontrado más pruebas de saurópodos de este tamaño, y el descubridor, Edward Cope, había cometido errores tipográficos antes, es probable que haya sido una sobreestimación extrema. A partir de 2018, Argentinosaurus y Patagotitan son considerados por los paleontólogos como los dinosaurios más grandes conocidos a partir de restos razonables.
El dinosaurio carnívoro más grande era Spinosaurus , alcanzando una longitud de 12.6 a 18 metros (41 a 59 pies) y un peso de 7-20.9 toneladas (7.7-23 toneladas cortas). Otros grandes terópodos carnívoros incluyen Giganotosaurus , Carcharodontosaurus y Tyrannosaurus . Therizinosaurus y Deinocheirus estaban entre los terópodos más altos. El Ornithischiandinosaur más grande fue probablemente el hadrosáurido Shantungosaurus que medía 16 metros (52 pies) y pesaba alrededor de 13 toneladas (29,000 lb).
El dinosaurio más pequeño que se conoce es el colibrí abeja, con una longitud de solo 5 cm (2.0 in) y una masa de alrededor de 1.8 g (0.063 oz). Los más pequeños conocidos como no-avialandinosaurios tenían aproximadamente el tamaño de las palomas y eran los terópodos más estrechamente relacionados con las aves. Por ejemplo, Anchiornis huxleyi es actualmente el dinosaurio no aviano más pequeño descrito de un espécimen adulto, con un peso estimado de 110 gramos y una longitud esquelética total de 34 cm (1.12 pies). Los dinosaurios herbívoros no aviales más pequeños incluían Microceratus y Wannanosaurus , de aproximadamente 60 cm (2.0 pies) de largo cada uno.
Comportamiento

Muchas aves modernas son muy sociables, a menudo se encuentran viviendo en bandadas. Existe un acuerdo general de que algunos comportamientos que son comunes en las aves, así como en los cocodrilos (parientes vivos más cercanos de las aves), también eran comunes entre los grupos de dinosaurios extintos. Las interpretaciones del comportamiento de las especies fósiles generalmente se basan en la postura de los esqueletos y su hábitat, las simulaciones informáticas de su biomecánica y las comparaciones con animales modernos en nichos ecológicos similares.
La primera evidencia potencial de pastoreo o reunión como un comportamiento generalizado común a muchos grupos de dinosaurios además de aves fue el descubrimiento de 1878 de 31 Iguanodon bernissartensis, ornitisquios que se pensaba que habían perecido juntos en Bernissart, Bélgica, después de haber caído en un sumidero profundo e inundado y haberse ahogado. Otros sitios de muerte en masa se han descubierto posteriormente. Esos, junto con múltiples caminos, sugieren que el comportamiento gregario era común en muchas especies tempranas de dinosaurios. Los senderos de cientos o incluso miles de herbívoros indican que los patos-aguja (hadrosáuridos) pueden haberse movido en grandes manadas, como el bisonte americano o el gacebo africano. Las huellas de saurópodos documentan que estos animales viajaron en grupos compuestos de varias especies diferentes, al menos en Oxfordshire, Inglaterra, aunque no hay evidencia de estructuras de rebaños específicas. Congregándose en rebaños puede haber evolucionado para la defensa, con fines migratorios, o para proporcionar protección a los jóvenes. Hay evidencia de que muchos tipos de dinosaurios de crecimiento lento, incluyendo varios terópodos, saurópodos, anquilosaurios, ornitópodos y ceratópsidos formaron agregaciones de individuos inmaduros. Un ejemplo es un sitio en Mongolia Interior que ha arrojado los restos de más de 20 Sinornithomimus , de uno a siete años. Este conjunto se interpreta como un grupo social que quedó atrapado en el barro. La interpretación de los dinosaurios como gregarios también se ha extendido a representar a los terópodos carnívoros como cazadores de paquetes que trabajan juntos para derribar presas grandes. Sin embargo, este estilo de vida es poco común entre aves modernas, cocodrilos y otros reptiles, y la evidencia tafonómica que sugiere la caza de manadas tipo mamífero en los terópodos como Deinonychus y Allosaurus también puede interpretarse como el resultado de disputas fatales entre animales de alimentación, como se ve en muchos depredadores de diapsidos modernos.

Las crestas y volantes de algunos dinosaurios, como los marginocefalianos, terópodos y lambeosaurinos, pueden haber sido demasiado frágiles para ser utilizados para la defensa activa, por lo que probablemente se usaron para exhibiciones sexuales o agresivas, aunque se sabe poco sobre apareamiento de dinosaurios y territorialismo. Las heridas en la cabeza causadas por las picaduras sugieren que los terópodos, al menos, se enfrentaron de forma activa y agresiva.
Desde el punto de vista del comportamiento, uno de los fósiles de dinosaurios más valiosos fue descubierto en el desierto de Gobi en 1971. Incluía un Velociraptor atacando a un Protoceratops , proporcionando evidencia de que los dinosaurios sí se atacaban entre sí. La evidencia adicional para atacar presas vivas es la cola parcialmente curada de un Edmontosaurus , un dinosaurio hadrosáurido; la cola está dañada de tal manera que muestra que el animal fue mordido por un tiranosaurio pero sobrevivió. El canibalismo entre algunas especies de dinosaurios fue confirmado por marcas de dientes encontradas en Madagascar en 2003, que involucraron al terópodo Majungasaurus .
Las comparaciones entre los anillos esclerales de los dinosaurios y las aves y reptiles modernos se han utilizado para inferir los patrones de actividad diaria de los dinosaurios. Aunque se ha sugerido que la mayoría de los dinosaurios estuvieron activos durante el día, estas comparaciones han demostrado que los pequeños dinosaurios depredadores como los dromeosáuridos, Juravenator y Megapnosaurus eran probablemente nocturnos. Los dinosaurios herbívoros y omnívoros grandes y medianos, como los ceratópsidos, sauropodomorfos, hadrosáuridos y ornitomimatosos, pueden haber sido catetrales, activos durante intervalos cortos durante todo el día, aunque se infirió que el pequeño ornitisquio Agilisaurus era diurno.
Con base en la evidencia fósil actual de los dinosaurios como el Oryctodromeus , algunas especies ornitisquias parecen haber llevado un estilo de vida parcialmente fossorial. Muchas aves modernas son arbóreas (trepar a los árboles), y esto también fue cierto para muchas aves mesozoicas, especialmente las enantiornitinas. Mientras que algunas especies tempranas de aves también pueden haber sido ya arbóreas (incluyendo dromeosáuridos como Microraptor) la mayoría de los dinosaurios no aviales parecen haberse basado en la locomoción terrestre. Una buena comprensión de cómo se movían los dinosaurios en el suelo es clave para los modelos de comportamiento de los dinosaurios; la ciencia de la biomecánica, iniciada por Robert McNeill Alexander, ha proporcionado una visión significativa en esta área. Por ejemplo, los estudios de las fuerzas ejercidas por los músculos y la gravedad sobre la estructura esquelética de los dinosaurios han investigado qué tan rápido podrían correr los dinosaurios, si los diplodócidos podrían crear auges sónicos a través del chasquido de la cola, y si los saurópodos podrían flotar.
Comunicación

Se sabe que las aves modernas se comunican usando señales visuales y auditivas, y la gran diversidad de estructuras de visualización entre grupos de dinosaurios fósiles, como cuernos, adornos, crestas, velas y plumas, sugiere que la comunicación visual siempre ha sido importante en la biología de los dinosaurios. Reconstrucción del color del plumaje de Anchiornis huxleyi, sugieren la importancia del color en la comunicación visual en dinosaurios no aviares. La evolución de la vocalización de los dinosaurios es menos segura. El paleontólogo Phil Senter sugiere que los dinosaurios no aviares se basaban principalmente en pantallas visuales y posiblemente sonidos acústicos no vocales como silbidos, aplastamiento de mandíbulas o aplausos, salpicaduras y golpes en las alas (posible en los dinosaurios maniraptoranos alados). Afirma que era improbable que fueran capaces de vocalizar ya que sus parientes más cercanos, cocodrilos y pájaros usan diferentes medios para vocalizar, el primero a través de la laringe y el último a través de la única siringe, sugiriendo que evolucionaron independientemente y que su ancestro común estaba mudo.
Los restos más antiguos de una siringe, que tiene suficiente contenido mineral para la fosilización, se encontraron en un espécimen del tipo pato Vegavis iaai data de hace 69-66 millones de años, y es poco probable que este órgano haya existido en los dinosaurios no aviares. Sin embargo, a diferencia de Senter, los investigadores sugirieron que los dinosaurios podían vocalizar y que el sistema vocal de aves basado en syrinx evolucionó a partir de uno basado en la laringe, en lugar de que los dos sistemas evolucionaran de forma independiente. Un estudio de 2016 sugiere que los dinosaurios produjeron vocalizaciones de boca cerrada, como el arrullo, que ocurren tanto en cocodrilos y aves como en otros reptiles. Dichas vocalizaciones evolucionaron de forma independiente en los arcosaurios existentes en numerosas ocasiones, después de aumentos en el tamaño corporal. Se ha sugerido que las crestas de los Lambeosaurini y las cámaras nasales de anquilosaurios funcionan en resonancia vocal.
Biología reproductiva

Todos los dinosaurios ponen huevos amnióticos con conchas duras hechas principalmente de carbonato de calcio. Los huevos generalmente se ponen en un nido. La mayoría de las especies crean nidos algo elaborados, que pueden ser copas, cúpulas, placas, raspaduras de camas, montículos o madrigueras. Algunas especies de aves modernas no tienen nidos; el arao común que anida en un acantilado pone sus huevos en la roca desnuda, y los pingüinos emperador macho mantienen los huevos entre su cuerpo y los pies. Las aves primitivas y muchos dinosaurios no aviales a menudo ponen huevos en nidos comunales, y los machos principalmente incuban los huevos. Mientras que las aves modernas solo tienen un oviducto funcional y ponen un huevo a la vez, las aves más primitivas y los dinosaurios tenían dos oviductos, como los cocodrilos. Algunos dinosaurios no aviales, como Troodon, exhibió la colocación iterativa, donde el adulto podría poner un par de huevos cada uno o dos días, y luego aseguró la eclosión simultánea al retrasar la cría hasta que todos los huevos fueron colocados.
Cuando ponen huevos, las hembras desarrollan un tipo especial de hueso entre el hueso externo duro y la médula de sus extremidades. Este hueso medular, que es rico en calcio, se usa para hacer cáscaras de huevo. Un descubrimiento de características en un esqueleto de Tyrannosaurus rex proporcionó evidencia de hueso medular en dinosaurios extintos y, por primera vez, permitió a los paleontólogos establecer el sexo de un espécimen fósil de dinosaurio. La investigación adicional ha encontrado hueso medular en el carnosaur Allosaurus y el ornitópodo Tenontosaurus . Porque la línea de dinosaurios que incluye Allosaurus y Tyrannosaurus divergieron de la línea que llevaba a Tenontosaurus muy temprano en la evolución de los dinosaurios, esto sugiere que la producción de tejido medular es una característica general de todos los dinosaurios.

Otro rasgo generalizado entre las aves modernas (pero véase más adelante en lo que respecta a los grupos fósiles y megapodes existentes) es el cuidado parental de los jóvenes después de la eclosión. El descubrimiento de Jack Horner en 1978 de un nidalen Maiasaura ("buena madre lagarto") en Montana demostró que el cuidado parental continuó mucho después del nacimiento entre los ornitópodos. En 1993 se descubrió un ejemplar del oviraptorido mongol Citipati osmolskae en una posición de cría parecida a la de una gallina, lo que puede indicar que habían comenzado a usar una capa aislante de plumas para mantener los huevos calientes. Un embrión de dinosaurio (perteneciente al prosaurópodo Massospondylus) fue encontrado sin dientes, lo que indica que se requería cierto cuidado parental para alimentar a los dinosaurios jóvenes. Las vías también han confirmado el comportamiento de los padres entre los ornitópodos de la isla de Skye, en el noroeste de Escocia.
Sin embargo, existe amplia evidencia de supremacía entre muchas especies de dinosaurios, particularmente terópodos. Por ejemplo, se ha demostrado abundantemente que las aves no ornituromorfas han tenido tasas de crecimiento lento, comportamiento de enterramiento de huevos similar a un megapode y la capacidad de volar poco después del nacimiento. Tanto Tyrannosaurus rex como Troodon formosus muestran juveniles con supremacía evidente y probablemente ocupando diferentes nichos ecológicos que los adultos. La superprecocialidad se ha inferido para los saurópodos.
Fisiología
Debido a que tanto los cocodrilos modernos como las aves tienen corazones de cuatro cámaras (aunque modificados en cocodrílidos), es probable que este sea un rasgo compartido por todos los arcosaurios, incluidos todos los dinosaurios. Si bien todas las aves modernas tienen metabolismos elevados y son de "sangre caliente" (endotérmica), desde la década de 1960 se ha mantenido un vigoroso debate sobre cuán atrás en el linaje de los dinosaurios se extiende este rasgo. Los científicos no están de acuerdo en cuanto a si los dinosaurios no aviares eran endotérmicos, ectotérmicos o una combinación de ambos.
Después de que se descubrieron los dinosaurios no aviares, los paleontólogos primero postularon que eran ectotérmicos. Esta supuesta "sangre fría" se utilizó para dar a entender que los antiguos dinosaurios eran organismos lentos y lentos, aunque muchos reptiles modernos son rápidos y ligeros a pesar de depender de fuentes externas de calor para regular la temperatura de su cuerpo. La idea de los dinosaurios como ectotérmicos y lentos siguió siendo una visión prevalente hasta que Robert T. "Bob" Bakker, uno de los primeros defensores de la endotermia de los dinosaurios, publicó un influyente artículo sobre el tema en 1968.
La evidencia moderna indica que incluso los dinosaurios no aviar y las aves prosperaron en climas templados más fríos, y que al menos algunas especies tempranas deben haber regulado su temperatura corporal por medios biológicos internos (ayudados por la masa de los animales en especies grandes y plumas u otras cubiertas corporales en especies más pequeñas). La evidencia de endotermia en los dinosaurios mesozoicos incluye el descubrimiento de dinosaurios polares en Australia y la Antártida, así como el análisis de las estructuras de los vasos sanguíneos dentro de los huesos fósiles que son típicos de endotermos. Continúa el debate científico sobre las formas específicas en que evolucionó la regulación de la temperatura de los dinosaurios.

En los dinosaurios saurisquios, los metabolismos más elevados se vieron respaldados por la evolución del sistema respiratorio aviar, que se caracteriza por un sistema extensivo de sacos de aire que extendieron los pulmones e invadieron muchos de los huesos del esqueleto, haciéndolos huecos. Sistemas respiratorios de estilo temprano aviar con los sacos aéreos pueden haber sido capaces de mantener niveles de actividad más elevados que los de mamíferos de tamaño y construcción similares. Además de proporcionar un suministro de oxígeno muy eficiente, el flujo de aire rápido habría sido un mecanismo de enfriamiento efectivo, que es esencial para los animales que son activos pero demasiado grandes para eliminar todo el exceso de calor a través de su piel.
Al igual que otros reptiles, los dinosaurios son principalmente uricotélicos, es decir, sus riñones extraen desechos nitrogenados de su torrente sanguíneo y los excretan como ácido úrico en lugar de urea o amoníaco a través de los uréteres en el intestino. En la mayoría de las especies vivas, el ácido úrico se excreta junto con las heces como un desecho semisólido. Sin embargo, al menos algunas aves modernas (como los colibríes) pueden ser facultativamente amonotelóticas, excretando la mayoría de los desechos nitrogenados en forma de amoniaco. También excretan creatina, en lugar de creatinina como los mamíferos. Este material, así como la salida de los intestinos, emerge de la cloaca. Además, muchas especies regurgitan gránulos, y los gránulos fósiles que pueden haber venido de los dinosaurios se conocen desde hace mucho tiempo como el período Cretácico.
Origen de las aves
La posibilidad de que los dinosaurios fueran los antepasados de las aves fue sugerida por primera vez en 1868 por Thomas Henry Huxley. Después del trabajo de Gerhard Heilmann a principios del siglo XX, la teoría de las aves como descendientes de dinosaurios fue abandonada a favor de la idea de que fueran descendientes de los tecodondos generalizados, y la evidencia clave es la supuesta falta de clavículas en los dinosaurios. Sin embargo, como demostraron los descubrimientos posteriores, las clavículas (o un único esqueleto fusionado, que se deriva de clavículas separadas) en realidad no estaban ausentes; se habían encontrado ya en 1924 en Oviraptor, pero identificado erróneamente como una interclavícula. En la década de 1970, John Ostrom renunció a la teoría de los dinosaurios-pájaros, que ganó impulso en las próximas décadas con el advenimiento del análisis cladístico y un gran aumento en el descubrimiento de pequeños terópodos y aves tempranas. De particular interés han sido los fósiles de la Formación Yixian, donde se han encontrado una gran variedad de terópodos y aves tempranas, a menudo con plumas de algún tipo. Las aves comparten más de cien características anatómicas distintas con los dinosaurios terópodos, que ahora se acepta generalmente como sus parientes más cercanos. Están estrechamente relacionados con los celurosaurios maniraptores. Una minoría de científicos, en particular Alan Feduccia y Larry Martin, han propuesto otros caminos evolutivos, incluidas versiones revisadas de la propuesta basil de los arcosaurios de Heilmann,
Plumas

Las plumas son una de las características más reconocibles de las aves modernas, y un rasgo que fue compartido por todos los demás grupos de dinosaurios. Con base en la distribución actual de la evidencia fósil, parece que las plumas eran un rasgo dinosaurio ancestral, aunque se las pudo haber perdido selectivamente en algunas especies. Se ha descubierto evidencia fósil directa de plumas o estructuras similares a plumas en una gran variedad de especies en muchos grupos de dinosaurios no aviares, tanto entre saurios como ornitisquios. Las estructuras simples, ramificadas, parecidas a plumas se conocen de los heterodontosáuridos, los primitivos neorritisquios y terópodos, y los ceratópsidos primitivos. La evidencia de verdaderas plumas con alas similares a las plumas de vuelo de las aves modernas se ha encontrado solo en el subgrupo de terópodos Maniraptora, que incluye oviraptorosaurios, troodóntidos, dromeosáuridos, y pájaros. También se han encontrado estructuras en forma de pluma conocidas como picnofibras en los pterosaurios, lo que sugiere la posibilidad de que los filamentos parecidos a plumas hayan sido comunes en el linaje de las aves y hayan evolucionado antes de la aparición de los propios dinosaurios. La investigación sobre la genética de los caimanes americanos también ha revelado que los escudos cocodrilos poseen plumas queratinas durante el desarrollo embrionario, pero estas queratinas no son expresadas por los animales antes de la eclosión.
Archaeopteryx fue el primer fósil encontrado que reveló una posible conexión entre los dinosaurios y las aves. Se considera un fósil de transición, ya que muestra características de ambos grupos. Expuesto a la luz solo dos años después del influyente libro de Darwin, The Origin of Species , su descubrimiento estimuló el debate naciente entre los defensores de la biología evolutiva y el creacionismo. Esta madriguera es tan similar a un dinosaurio, sin una impresión clara de plumas en la roca circundante, al menos un espécimen fue confundido con Compsognathus. Desde la década de 1990, se han encontrado una serie de dinosaurios emplumados adicionales, que proporcionan evidencia aún más sólida de la estrecha relación entre los dinosaurios y las aves modernas. La mayoría de estos especímenes fueron desenterrados en la lagerstätte de la Formación Yixian, Liaoning, noreste de China, que fue parte de un continente insular durante el Cretácico. Aunque se han encontrado plumas en solo unos pocos lugares, es posible que los dinosaurios no avianos en otras partes del mundo también hayan sido emplumados. La falta de evidencia fósil generalizada para los dinosaurios emplumados no aviares puede deberse a que las características delicadas, como la piel y las plumas, a menudo no se preservan mediante la fosilización y, por lo tanto, están ausentes del registro fósil.
La descripción de los dinosaurios emplumados no ha estado libre de controversia; quizás los críticos más vocales han sido Alan Feduccia y Theagarten Lingham-Soliar, que han propuesto que algunos supuestos fósiles parecidos a plumas son el resultado de la descomposición de la fibra de colágeno que subyace a la piel de los dinosaurios, y que los dinosaurios maniraptoranos con plumas en realidad dinosaurios, pero convergentes con los dinosaurios. Sin embargo, sus puntos de vista en su mayoría no han sido aceptados por otros investigadores, hasta el punto de que la naturaleza científica de las propuestas de Feduccia ha sido cuestionada.
En 2016, se informó que una cola de dinosaurio con plumas se había encontrado encerrada en ámbar. El fósil tiene alrededor de 99 millones de años.
Esqueleto
Debido a que las plumas a menudo se asocian con las aves, los dinosaurios emplumados a menudo se promocionan como el eslabón perdido entre las aves y los dinosaurios. Sin embargo, las múltiples características esqueléticas también compartidas por los dos grupos representan otra línea de evidencia importante para los paleontólogos. Las áreas del esqueleto con similitudes importantes incluyen el cuello, el pubis, la muñeca (carpo semilunar), el brazo y la cintura pectoral, la fúrcula (esternón) y el esternón. La comparación de los esqueletos de aves y dinosaurios a través del análisis cladístico fortalece el caso del enlace.
Anatomía suave

Los grandes dinosaurios carnívoros tenían un complejo sistema de sacos de aire similares a los que se encuentran en las aves modernas, según una investigación de 2005 dirigida por Patrick M. O'Connor. Los pulmones de los dinosaurios terópodos (carnívoros que caminaban sobre dos patas y tenían patas de pájaro) probablemente bombeaban aire dentro de los sacos huecos de sus esqueletos, como es el caso de las aves. "Lo que una vez se consideró formalmente exclusivo de las aves estuvo presente de alguna forma en los ancestros de las aves", dijo O'Connor. En 2008, los científicos describieron Aerosteon riocoloradensis , cuyo esqueleto proporciona la evidencia más fuerte hasta la fecha de un dinosaurio con un sistema de respiración similar a un ave. La tomografía computarizada de huesos fósiles de Aerosteon reveló evidencia de la existencia de bolsas de aire dentro de la cavidad del cuerpo del animal.
Evidencia conductual
Los fósiles de los troodontes Mei y Sinornithoides demuestran que algunos dinosaurios dormían con sus cabezas metidas bajo sus brazos. Este comportamiento, que puede haber ayudado a mantener la cabeza caliente, también es característico de las aves modernas. Varios especímenes deinonchosaurio y oviraptorosaurio también se han encontrado preservados en la parte superior de sus nidos, probablemente meditando en forma de pájaro. La relación entre el volumen del huevo y la masa corporal de los adultos entre estos dinosaurios sugiere que los huevos fueron primordialmente medidos por el macho, y que los juveniles eran altamente precoces, similares a muchas aves modernas que viven en el suelo.
Se sabe que algunos dinosaurios usaron piedras de molleja como las aves modernas. Estas piedras son tragadas por los animales para ayudar a la digestión y descomponer los alimentos y las fibras duras una vez que ingresan al estómago. Cuando se encuentran en asociación con fósiles, las mollejas se llaman gastrolitos.
Extinción de grupos principales
El descubrimiento de que las aves son un tipo de dinosaurio mostró que los dinosaurios en general no están, de hecho, extintos como se afirma comúnmente. Sin embargo, todos los dinosaurios no aviares, que se estima que fueron 628-1078 especies, así como muchos grupos de aves se extinguieron repentinamente hace aproximadamente 66 millones de años. Se ha sugerido que debido a que pequeños mamíferos, squamata y aves ocupaban los nichos ecológicos adecuados para un tamaño corporal pequeño, los dinosaurios no aviares nunca desarrollaron una variada fauna de especies de cuerpo pequeño, lo que provocó su caída cuando los tetrápodos terrestres de gran tamaño fueron afectados por el evento de extinción masiva. Muchos otros grupos de animales también se extinguieron en este momento, incluidos amonites (moluscos similares al nautilus), mosasaurios, plesiosaurios, pterosaurios y muchos grupos de mamíferos. Significativamente, los insectos no sufrieron una pérdida de población discernible, que los dejó disponibles como alimento para otros sobrevivientes. Esta extinción masiva se conoce como el evento de extinción Cretácico-Paleógeno. La naturaleza del evento que causó esta extinción masiva ha sido ampliamente estudiada desde la década de 1970; en la actualidad, varias teorías relacionadas son apoyadas por los paleontólogos. Aunque el consenso es que un evento de impacto fue la causa principal de la extinción de los dinosaurios, algunos científicos citan otras posibles causas, o apoyan la idea de que una confluencia de varios factores fue responsable de la desaparición repentina de los dinosaurios del registro fósil. varias teorías relacionadas son apoyadas por los paleontólogos. Aunque el consenso es que un evento de impacto fue la causa principal de la extinción de los dinosaurios, algunos científicos citan otras posibles causas, o apoyan la idea de que una confluencia de varios factores fue responsable de la desaparición repentina de los dinosaurios del registro fósil. varias teorías relacionadas son apoyadas por los paleontólogos. Aunque el consenso es que un evento de impacto fue la causa principal de la extinción de los dinosaurios, algunos científicos citan otras posibles causas, o apoyan la idea de que una confluencia de varios factores fue responsable de la desaparición repentina de los dinosaurios del registro fósil.
Evento de impacto

La teoría de la colisión de asteroides, que Walter Alvarez y sus colegas llamaron la atención en 1980, relaciona el evento de extinción al final del período Cretácico con un impacto bólido hace aproximadamente 66 millones de años. Alvarez et al.propuso que un aumento repentino en los niveles de iridio, registrado en todo el mundo en el estrato rocoso del período, era una evidencia directa del impacto. El grueso de la evidencia ahora sugiere que un bólido de 5 a 15 kilómetros (3.1 a 9.3 millas) de ancho golpeó en las cercanías de la Península de Yucatán (en el sureste de México), creando el cráter Chicxulub de aproximadamente 180 km (110 millas) y desencadenando la masa extinción. Los científicos no están seguros de si los dinosaurios prosperaron o declinaron antes del evento de impacto. Algunos científicos proponen que el impacto del meteorito causó una caída larga y antinatural en la temperatura atmosférica de la Tierra, mientras que otros afirman que en su lugar habría creado una ola de calor inusual. El consenso entre los científicos que respaldan esta teoría es que el impacto causó extinciones tanto directamente (por el calor del impacto del meteorito) como indirectamente (a través de un enfriamiento mundial provocado cuando la materia expulsada del cráter de impacto reflejaba la radiación térmica del sol). Aunque la velocidad de extinción no se puede deducir solo del registro fósil, varios modelos sugieren que la extinción fue extremadamente rápida, ya que se redujo a horas en lugar de años.
Trampas de Deccan
Antes de 2000, los argumentos de que las inundaciones de Deccan Traps basaltos causaban la extinción solían relacionarse con la visión de que la extinción era gradual, ya que los eventos de inundación de basalto se iniciaron hace unos 68 millones de años y duraron más de 2 millones de años. Sin embargo, hay evidencia de que dos tercios de las trampas de Deccan se crearon en solo 1 millón de años hace 66 millones de años, por lo que estas erupciones habrían causado una extinción bastante rápida, posiblemente durante un período de miles de años, pero aún más que se esperaría de un solo evento de impacto.
Las trampas de Deccan en la India podrían haber causado la extinción a través de varios mecanismos, incluida la liberación al aire de polvo y aerosoles sulfúricos, que podrían haber bloqueado la luz solar y, por lo tanto, reducido la fotosíntesis en las plantas. Además, el volcanismo de Deccan Trap podría haber producido emisiones de dióxido de carbono, lo que habría incrementado el efecto invernadero cuando el polvo y los aerosoles se despejaron de la atmósfera. Antes de la extinción masiva de los dinosaurios, la liberación de gases volcánicos durante la formación de las trampas Deccan "contribuyó a un calentamiento global aparentemente masivo. Algunos datos apuntan a un aumento promedio de la temperatura de 8 ° C (14 ° F) en la última mitad millones de años antes del impacto [en Chicxulub] ".
En los años en que la teoría de Deccan Traps estaba vinculada a una extinción más lenta, Luis Álvarez (quien murió en 1988) respondió que los paleontólogos estaban siendo engañados por la escasez de datos. Si bien su afirmación al principio no fue bien recibida, posteriores estudios de campo intensivos de lechos fósiles le dieron peso a su afirmación. Eventualmente, la mayoría de los paleontólogos comenzaron a aceptar la idea de que las extinciones masivas al final del Cretácico se debieron en gran parte o al menos en parte a un impacto masivo en la Tierra. Sin embargo, incluso Walter Alvarez ha reconocido que hubo otros cambios importantes en la Tierra incluso antes del impacto, como una caída en el nivel del mar y erupciones volcánicas masivas que produjeron las trampas Deccan de la India, y que pueden haber contribuido a la extinción.
Posibles sobrevivientes del Paleoceno
Los restos de dinosaurios no aviares se encuentran ocasionalmente por encima del límite Cretácico-Paleógeno. En 2001, los paleontólogos Zielinski y Budahn informaron el descubrimiento de un único fósil de hueso de la pata hadrosaurio en la cuenca de San Juan, Nuevo México, y lo describieron como evidencia de dinosaurios del Paleoceno. La formación en la que se descubrió el hueso data de la época del Paleoceno temprano, hace aproximadamente 64,5 millones de años. Si el hueso no se volvió a depositar en ese estrato por acción de intemperie, proporcionaría evidencia de que algunas poblaciones de dinosaurios pueden haber sobrevivido al menos medio millón de años en la Era Cenozoica. Otra evidencia incluye el hallazgo de restos de dinosaurios en la Formación Hell Creek hasta 1,3 m (51 in) por encima del límite Cretácico-Paleógeno, lo que representa 40 000 años de tiempo transcurrido. Informes similares han llegado de otras partes del mundo, incluida China. Muchos científicos, sin embargo, descartaron los supuestos dinosaurios del Paleoceno como reelaborados, es decir, eliminados de sus lugares originales y luego vueltos a enterrar en sedimentos mucho más tardíos. La datación directa de los propios huesos ha respaldado la fecha posterior, con los métodos de datación U-Pb que dan como resultado una edad precisa de 64.8 ± 0.9 millones de años atrás. Si es correcto, la presencia de un puñado de dinosaurios en el Paleoceno temprano no cambiaría los hechos subyacentes de la extinción.
Historia del estudio
Los fósiles de dinosaurios se conocen desde hace milenios, aunque su verdadera naturaleza no fue reconocida. Los chinos, cuya palabra moderna para dinosaurio es kǒnglóng (恐龍, o "terrible dragón"), los consideraron como huesos de dragón y los documentaron como tales. Por ejemplo, Hua Yang Guo Zhi , un libro escrito por Chang Qu durante la dinastía Jin occidental (265-316), informó sobre el descubrimiento de huesos de dragón en Wucheng en la provincia de Sichuan. Los aldeanos en el centro de China han desenterrado durante mucho tiempo huesos de dragón fosilizados para su uso en medicinas tradicionales, una práctica que continúa en la actualidad. En Europa, se creía que los fósiles de dinosaurios eran los restos de gigantes y otras criaturas bíblicas.
Las descripciones académicas de lo que ahora se reconocería como huesos de dinosaurios aparecieron por primera vez a fines del siglo XVII en Inglaterra. Parte de un hueso, que ahora se conoce como el fémur de un Megalosaurus , fue recuperado de una cantera de piedra caliza en Cornwell cerca de Chipping Norton, Oxfordshire, en 1676. El fragmento fue enviado a Robert Plot, profesor de Química en la Universidad de Oxford. primer curador del Museo Ashmolean, quien publicó una descripción en su Historia Natural de Oxfordshireen 1677. Identificó correctamente el hueso como la extremidad inferior del fémur de un animal grande, y reconoció que era demasiado grande para pertenecer a cualquier especie conocida. Por lo tanto, concluyó que era el hueso del muslo de un humano gigante similar a los mencionados en la Biblia. En 1699, Edward Lhuyd, amigo de Sir Isaac Newton, fue el responsable del primer tratamiento científico publicado de lo que ahora sería reconocido como un dinosaurio cuando describió y nombró un diente de saurópodo, "Rutellum implicatum", que se había encontrado en Caswell , cerca de Witney, Oxfordshire.

Entre 1815 y 1824, el reverendo William Buckland, profesor de geología en Oxford, recolectó más huesos fosilizados de Megalosaurus y se convirtió en la primera persona en describir un dinosaurio en una revista científica. El segundo género de dinosaurio identificado, Iguanodon , fue descubierto en 1822 por Mary Ann Mantell, la esposa del geólogo inglés Gideon Mantell. Gideon Mantell reconoció similitudes entre sus fósiles y los huesos de las iguanas modernas. Publicó sus hallazgos en 1825.
El estudio de estos "grandes lagartos fósiles" pronto se convirtió en un gran interés para los científicos europeos y estadounidenses, y en 1842 el paleontólogo inglés Richard Owen acuñó el término "dinosaurio". Reconoció que los restos que se habían encontrado hasta ahora, Iguanodon , Megalosaurus e Hylaeosaurus , compartían una serie de características distintivas, por lo que decidieron presentarlos como un grupo taxonómico distinto. Con el respaldo del Príncipe Alberto, el esposo de la Reina Victoria, Owen estableció el Museo de Historia Natural de Londres para exhibir la colección nacional de fósiles de dinosaurios y otras exhibiciones biológicas y geológicas.
En 1858, William Parker Foulke descubrió el primer dinosaurio estadounidense conocido, en pozos de marga en la pequeña ciudad de Haddonfield, Nueva Jersey. (Aunque los fósiles se habían encontrado antes, su naturaleza no se había discernido correctamente.) La criatura se llamó Hadrosaurus foulkii . Fue un descubrimiento extremadamente importante: Hadrosaurus fue uno de los primeros esqueletos de dinosaurios casi completos encontrados (el primero fue en 1834, en Maidstone, Inglaterra), y era claramente una criatura bípeda. Este fue un descubrimiento revolucionario ya que, hasta ese momento, la mayoría de los científicos creían que los dinosaurios caminaban a cuatro patas, como otros lagartos. Los descubrimientos de Foulke desató una oleada de manía de dinosaurios en los Estados Unidos.


La manía de los dinosaurios fue ejemplificada por la feroz rivalidad entre Edward Drinker Cope y Othniel Charles Marsh, quienes corrieron para ser los primeros en encontrar nuevos dinosaurios en lo que llegó a conocerse como las Guerras de los Huesos. La disputa probablemente se originó cuando Marsh señaló públicamente que la reconstrucción de Cope de un Elasmosaurio el esqueleto era defectuoso: Cope había colocado inadvertidamente la cabeza del plesiosaurio en lo que debería haber sido el final de la cola del animal. La pelea entre los dos científicos duró más de 30 años, terminando en 1897 cuando Cope murió después de gastar toda su fortuna en la caza de dinosaurios. Marsh 'ganó' el concurso principalmente porque fue mejor financiado a través de una relación con el Servicio Geológico de los Estados Unidos. Desafortunadamente, muchos especímenes valiosos de dinosaurios fueron dañados o destruidos debido a los métodos rudos de la pareja: por ejemplo, sus excavadores a menudo usaban dinamita para desenterrar huesos (un método que a los paleontólogos les parecería espantoso). A pesar de sus métodos no refinados, las contribuciones de Cope y Marsh a la paleontología fueron enormes: Marsh descubrió 86 nuevas especies de dinosaurios y Cope descubrió 56, un total de 142 nuevas especies. Capa pluvial'
Después de 1897, la búsqueda de fósiles de dinosaurios se extendió a todos los continentes, incluida la Antártida. El primer dinosaurio antártico que se descubrió, el anquilosaurio Antarctopelta oliveroi , se encontró en la isla James Ross en 1986, aunque fue en 1994 antes de que una especie antártica, el terópodo Cryolophosaurus ellioti , fuera formalmente nombrada y descrita en una revista científica.
Los "puntos calientes" actuales de los dinosaurios incluyen el sur de Sudamérica (especialmente Argentina) y China. China en particular ha producido muchos ejemplares excepcionales de dinosaurios emplumados debido a la geología única de sus capas de dinosaurios, así como a un antiguo clima árido particularmente propicio para la fosilización.
"Renacimiento de los dinosaurios"

El campo de la investigación de dinosaurios ha disfrutado de un aumento en la actividad que comenzó en la década de 1970 y está en curso. Este fue provocada, en parte, por el descubrimiento de John Ostrom de Deinonychus, un depredador activo que puede haber sido de sangre caliente, en marcado contraste con la imagen de los dinosaurios que entonces prevalecía, de lentos y de sangre fría. La paleontología de vertebrados se ha convertido en una ciencia global. Nuevos descubrimientos de dinosaurios han sido realizados por paleontólogos que trabajan en regiones previamente inexplotadas, incluyendo India, Sudamérica, Madagascar, Antártida y, lo más importante, China (los dinosaurios emplumados increíblemente bien conservados en China han consolidado aún más el vínculo entre los dinosaurios y sus descendientes vivos , pájaros modernos). La aplicación generalizada de la cladística, que analiza rigurosamente las relaciones entre organismos biológicos, también ha demostrado ser tremendamente útil en la clasificación de los dinosaurios. El análisis cladístico, entre otras técnicas modernas, ayuda a compensar un registro fósil a menudo incompleto y fragmentario.
TABINITABEND
Tejido blando y ADN

Uno de los mejores ejemplos de impresiones de tejidos blandos en un dinosaurio fósil fue descubierto en Pietraroia, Italia. El descubrimiento fue reportado en 1998 y describió el espécimen de un pequeño y muy joven coturosaurio, Scipionyx samniticus . El fósil incluye partes de los intestinos, colon, hígado, músculos y tráquea de este dinosaurio inmaduro.
En la edición de marzo de 2005 de Science , la paleontóloga Mary Higby Schweitzer y su equipo anunciaron el descubrimiento de material flexible parecido a un tejido blando real dentro de un Tyrannosaurus rex de 68 millones de años de antigüedad. hueso de la pierna de la Formación Hell Creek en Montana. Después de la recuperación, el equipo de ciencia rehidrató el tejido. Cuando el hueso fosilizado se trató durante varias semanas para eliminar el contenido mineral de la cavidad de médula ósea fosilizada (un proceso llamado desmineralización), Schweitzer encontró evidencia de estructuras intactas, como vasos sanguíneos, matriz ósea y tejido conectivo (fibras óseas). El escrutinio bajo el microscopio reveló además que el tejido blando del dinosaurio putativo había retenido estructuras finas (microestructuras) incluso a nivel celular. La naturaleza exacta y la composición de este material, y las implicaciones del descubrimiento de Schweitzer, aún no están claras.
En 2009, un equipo que incluía a Schweitzer anunció que, utilizando una metodología aún más cuidadosa, habían duplicado sus resultados al encontrar tejidos blandos similares en un dinosaurio con pico de pato, Brachylophosaurus canadensis , encontrado en la Formación Judith River de Montana. Esto incluyó tejido aún más detallado, hasta células óseas preservadas que incluso parecen tener restos visibles de núcleos y lo que parecen ser glóbulos rojos. Entre otros materiales encontrados en el hueso se encontraba el colágeno, como en el hueso Tyrannosaurus . El tipo de colágeno que tiene un animal en sus huesos varía según su ADN y, en ambos casos, este colágeno era del mismo tipo que se encuentra en los pollos y avestruces modernos.
La extracción de ADN antiguo de fósiles de dinosaurios se ha informado en dos ocasiones distintas; Sin embargo, luego de una inspección adicional y revisión por pares, ninguno de estos informes podría ser confirmado. Sin embargo, se ha deducido un péptido funcional involucrado en la visión de un dinosaurio teórico usando métodos analíticos de reconstrucción filogenética en secuencias genéticas de especies modernas relacionadas, tales como reptiles y aves. Además, varias proteínas, incluida la hemoglobina, se han detectado supuestamente en fósiles de dinosaurios.
En 2015, los investigadores informaron que encontraron estructuras similares a las células sanguíneas y las fibras de colágeno, conservadas en los fósiles de hueso de seis especímenes de dinosaurios del Cretácico, que tienen aproximadamente 75 millones de años.


Representaciones culturales
Para los estándares humanos, los dinosaurios eran criaturas de apariencia fantástica y, a menudo, de un tamaño enorme. Como tales, han capturado la imaginación popular y se han convertido en una parte perdurable de la cultura humana. La entrada de la palabra "dinosaurio" en la lengua vernácula común refleja la importancia cultural de los animales: en inglés, "dinosaurio" se usa comúnmente para describir cualquier cosa que sea imprácticamente grande, obsoleta o destinada a la extinción.
El entusiasmo público por los dinosaurios se desarrolló por primera vez en la Inglaterra victoriana, donde en 1854, tres décadas después de las primeras descripciones científicas de restos de dinosaurios, se inauguró una colección de esculturas de dinosaurios realistas en el Crystal Palace Park de Londres. Los dinosaurios Crystal Palace demostraron ser tan populares que pronto se desarrolló un mercado fuerte en réplicas más pequeñas. En las décadas siguientes, las exhibiciones de dinosaurios se abrieron en parques y museos de todo el mundo, lo que garantiza que las sucesivas generaciones se presentarán a los animales de una manera inmersiva y emocionante. La popularidad duradera de los dinosaurios, a su vez, ha dado como resultado una financiación pública significativa para la ciencia de los dinosaurios, y con frecuencia ha estimulado nuevos descubrimientos. En los Estados Unidos, por ejemplo, la competencia entre los museos por la atención pública condujo directamente a las Guerras Óseas de los años 1880 y 1890,
La preocupación popular por los dinosaurios ha asegurado su aparición en la literatura, el cine y otros medios. Comenzando en 1852 con una mención pasajera en la Casa Sombría de Charles Dickens , los dinosaurios han aparecido en gran cantidad de obras ficticias. La novela de Julio de 1864 Viaje al centro de la tierra , el libro de 1912 de Sir Arthur Conan Doyle The Lost World , la icónica película de 1933 King Kong , Godzilla de 1954 y sus numerosas secuelas, la novela de 1990 más famosa Jurassic Park por Michael Crichtone y su adaptación cinematográfica de 1993 son solo algunos ejemplos notables de apariciones de dinosaurios en la ficción. Los autores de obras de no ficción de interés general sobre los dinosaurios, incluidos algunos paleontólogos prominentes, a menudo han tratado de utilizar los animales como una forma de educar a los lectores sobre la ciencia en general. Los dinosaurios son omnipresentes en la publicidad; Numerosas compañías han hecho referencia a los dinosaurios en publicidades impresas o televisadas, ya sea para vender sus propios productos o para caracterizar a sus rivales como lentos, tontos u obsoletos.
Obtenido de: https://en.wikipedia.org/wiki/Dinosaur