Reptil
Definición
Los reptiles son animales tetrápodos en la clase Reptilia , que comprende tortugas, cocodrilos, serpientes, anfisbánidos, lagartos, tuataras y sus parientes extintos. El estudio de estas órdenes tradicionales de reptiles, históricamente combinadas con las de los anfibios modernos, se llama herpetología.Debido a que algunos reptiles están más estrechamente relacionados con las aves que con otros reptiles (por ejemplo, los cocodrilos están más estrechamente relacionados con las aves que con los lagartos), los grupos tradicionales de "reptiles" enumerados anteriormente no constituyen un grupo monofilético o clado (que consiste en todos los descendientes de un ancestro común). Por esta razón, muchos científicos modernos prefieren considerar a las aves como parte de Reptilia, convirtiendo así a Reptilia en una clase monofilética, que incluye a todos los diapsidos vivos.

En el sentido de las agujas del reloj desde arriba a la izquierda: Tortuga verde ( Chelonia mydas ),
Tuatara ( Sphenodon punctatus ), cocodrilo del Nilo
( Crocodylus niloticus ) y Sinaí agama ( Pseudotrapelus sinaitus ).
En el sentido de las agujas del reloj desde arriba a la izquierda: Tortuga verde ( Chelonia mydas ),
Tuatara ( Sphenodon punctatus ), cocodrilo del Nilo
( Crocodylus niloticus ) y Sinaí agama ( Pseudotrapelus sinaitus ).
Los proto-reptiles más antiguos conocidos se originaron hace alrededor de 312 millones de años durante el período Carbonífero, habiendo evolucionado a partir de tetrápodos reptiliomorfos avanzados que se adaptaron cada vez más a la vida en tierra firme. Algunos ejemplos tempranos incluyen Hylonomus y Casineria como lagarto . Además de los reptiles vivientes, existen muchos grupos diversos que ahora están extintos, en algunos casos debido a eventos de extinción en masa. En particular, el evento de extinción Cretácico-Paleógeno eliminó a los pterosaurios, plesiosaurios, ornitisquios y saurópodos, así como a muchas especies de terópodos, incluidos troodóntidos, dromeosáuridos, tiranosaurios y abelisáuridos, junto con muchos Crocodyliformes y escuamatos (p. Ej., Mosasauridos). .
Los reptiles no aviares modernos habitan todos los continentes excepto la Antártida, aunque algunas aves se encuentran en la periferia de la Antártida. Se reconocen varios subgrupos vivos: Testudines (tortugas y tortugas), 350 especies; Rhynchocephalia (tuatara de Nueva Zelanda), 1 especie; Squamata (lagartos, serpientes y lagartijas), más de 10,200 especies; Crocodilia (cocodrilos, gaviales, caimanes y lagartos), 24 especies; y Aves (aves), aproximadamente 10,000 especies.
Los reptiles son vertebrados tetrápodos, criaturas que tienen cuatro extremidades o, como las serpientes, descienden de antepasados de cuatro extremidades. A diferencia de los anfibios, los reptiles no tienen una etapa de larva acuática. La mayoría de los reptiles son ovíparos, aunque varias especies de escamosos son vivíparos, como lo fueron algunos clados acuáticos extintos: el feto se desarrolla dentro de la madre, contenido en una placenta en lugar de una cáscara de huevo. Como amniotas, los huevos de reptiles están rodeados por membranas de protección y transporte, que los adaptan a la reproducción en tierra firme. Muchas de las especies vivíparas alimentan a sus fetos a través de varias formas de placenta análogas a las de los mamíferos, y algunos brindan atención inicial para sus crías. Los reptiles existentes varían en tamaño desde un pequeño geco, Sphaerodactylus ariasae, que puede crecer hasta 17 mm (0,7 pulgadas) del cocodrilo de agua salada, Crocodylus porosus , que puede alcanzar los 6 m (19,7 pies) de longitud y pesar más de 1.000 kg (2.200 lb).
Clasificación
Historia de la investigación

En el siglo XIII, la categoría de reptiles fue reconocida en Europa como una mezcla de criaturas que ponen huevos, incluyendo "serpientes, diversos monstruos fantásticos, lagartos, anfibios variados y gusanos", según lo registrado por Vincent de Beauvais en su Espejo de naturaleza . En el siglo XVIII, los reptiles estaban, desde el comienzo de la clasificación, agrupados con los anfibios. Linneo, que trabaja en Suecia, donde la víbora común y la serpiente de hierba se encuentran a menudo cazando en el agua, incluía a todos los reptiles y anfibios de la clase "III - Amphibia" en su Systema Naturæ . Los términos "reptil" y "anfibio" eran en gran parte intercambiables, "reptil" (del latín repere, "arrastrarse") siendo preferido por los franceses. Josephus Nicolaus Laurenti fue el primero en utilizar formalmente el término "Reptilia" para una selección ampliada de reptiles y anfibios básicamente similar a la de Linnaeus. Hoy en día, los dos grupos todavía se tratan comúnmente bajo el mismo título que herptiles.

No fue sino hasta el comienzo del siglo XIX que quedó claro que los reptiles y anfibios son, de hecho, animales muy diferentes, y Pierre André Latreille erigió la clase Batracia (1825) para este último, dividiendo los tetrápodos en las cuatro clases familiares de reptiles, anfibios, aves y mamíferos. El anatomista británico Thomas Henry Huxley hizo popular la definición de Latreille y, junto con Richard Owen, amplió Reptilia para incluir los diversos "monstruos antediluvianos" fósiles, incluidos los dinosaurios y el dicinodon (sínapsis) parecido a los mamíferos. él ayudó a describir. Este no fue el único esquema de clasificación posible: en las conferencias de Hunterian pronunciadas en el Royal College of Surgeons en 1863, Huxley agrupó a los vertebrados en mamíferos, sauroides e ictioides (este último con peces y anfibios). Posteriormente propuso los nombres de Sauropsida e Ichthyopsida para los dos últimos grupos. En 1866, Haeckel demostró que los vertebrados podían dividirse en función de sus estrategias reproductivas, y que los reptiles, las aves y los mamíferos estaban unidos por el huevo amniótico.
Los términos "Sauropsida" ("caras de lagarto") y "Theropsida" ("caras de bestia") fueron utilizados nuevamente en 1916 por ES Goodrich para distinguir entre lagartos, aves y sus parientes por un lado (Sauropsida) y mamíferos y sus parientes extintos (Theropsida) por el otro. Goodrich apoyó esta división por la naturaleza de los corazones y vasos sanguíneos en cada grupo, y otras características, como la estructura del prosencéfalo. Según Goodrich, ambos linajes evolucionaron a partir de un grupo original anterior, Protosauria ("primeros lagartos") en el que incluyó algunos animales que hoy en día se consideran anfibios similares a reptiles, así como reptiles tempranos.
En 1956, DMS Watson observó que los primeros dos grupos divergían muy temprano en la historia de los reptiles, por lo que dividió Protosauria de Goodrich entre ellos. También reinterpretó a Sauropsida y Theropsida para excluir aves y mamíferos, respectivamente. Así, su Sauropsida incluía Procolophonia, Eosuchia, Millerosauria, Chelonia (tortugas), Squamata (lagartos y serpientes), Rhynchocephalia, Crocodilia, "thecodonts" (parachosa basal Archosauria), dinosaurios no aviares, pterosaurios, ictiosaurios y sauropterigios.
A fines del siglo XIX, se ofrecieron varias definiciones de Reptilia. Los rasgos enumerados por Lydekker en 1896, por ejemplo, incluyen un solo cóndilo occipital, una articulación de la mandíbula formada por el cuadrante y huesos articulares, y ciertas características de las vértebras. Los animales seleccionados por estas formulaciones, los amniotes distintos de los mamíferos y las aves, son todavía los considerados reptiles de la actualidad.

La división synapsid / sauropsid complementó otro enfoque, uno que divide los reptiles en cuatro subclases en función del número y la posición de fenestrae temporal, aberturas en los lados del cráneo detrás de los ojos. Esta clasificación fue iniciada por Henry Fairfield Osborn y elaborada y popularizada por la Paleontología Vertebrada clásica de Romer . Esas cuatro subclases fueron:
- Anapsida - no fenestra - cotylosaurs y Chelonia (tortugas y parientes)
- Synapsida - una baja fenestra - pelicosaurios y terápsidos (los "reptiles mamíferos")
- Euryapsida - un alto fenestra (por encima del postorbital y escamoso) - protorosaurios (pequeños reptiles de lagartija temprana) y los sauropterigios marinos e ictiosaurios, este último llamado Parapsida en el trabajo de Osborn.
- Diapsida - dos fenestrae - la mayoría de los reptiles, incluidos lagartos, serpientes, cocodrilos, dinosaurios y pterosaurios
La composición de Euryapsida era incierta. Los ictiosaurios fueron, a veces, considerados como surgidos independientemente de los otros euryapsidos, y se les dio el nombre más antiguo de Parapsida. Parapsida se descartó más tarde como grupo en su mayor parte (los ictiosaurios se clasificaron como incertae sedis o con Euryapsida). Sin embargo, cuatro (o tres si Euryapsida se fusiona con Diapsida) se mantuvieron más o menos universales para el trabajo no especializado a lo largo del siglo XX. Los investigadores recientes lo han abandonado en gran medida: en particular, se ha encontrado que la condición de la anapsid ocurre tan variablemente entre grupos no relacionados que ahora no se considera una distinción útil.
Filogenia y definición moderna

A principios del siglo XXI, los paleontólogos de vertebrados estaban empezando a adoptar la taxonomía filogenética, en la que todos los grupos se definen de forma tal que sean monofiléticos; es decir, los grupos incluyen todos los descendientes de un ancestro en particular. Los reptiles definidos históricamente son parafiléticos, ya que excluyen tanto a las aves como a los mamíferos. Estos evolucionaron respectivamente de dinosaurios y de terápsidos tempranos, que tradicionalmente se llamaron reptiles. Las aves están más relacionadas con los cocodrílidos que las últimas con el resto de los reptiles existentes. Colin Tudge escribió:
A pesar de las primeras propuestas para reemplazar la Reptilia parafilética con una Sauropsida monofilética, que incluye aves, ese término nunca se adoptó ampliamente o, cuando lo fue, no se aplicó de manera consistente. Cuando se usó Sauropsida, a menudo tenía el mismo contenido o incluso la misma definición que Reptilia. En 1988, Jacques Gauthier propuso una definición cladística de Reptilia como un grupo corona monofilético basado en nódulos que contiene tortugas, lagartos y serpientes, cocodrilos y aves, su ancestro común y todos sus descendientes. Debido a que la relación real de las tortugas con otros reptiles todavía no se entendía bien en este momento, la definición de Gauthier llegó a considerarse inadecuada.
Diversos científicos propusieron una variedad de otras definiciones en los años posteriores al artículo de Gauthier. La primera nueva definición, que intentó adherirse a los estándares del PhyloCode, fue publicada por Modesto y Anderson en 2004. Modesto y Anderson revisaron las muchas definiciones anteriores y propusieron una definición modificada, que pretendían conservar el contenido más tradicional de la grupo mientras lo mantiene estable y monofilético. Definieron a Reptilia como todos los amniotas más cercanos a Lacerta agilis y Crocodylus niloticus que a Homo sapiens.. Esta definición basada en el tronco es equivalente a la definición más común de Sauropsida, que Modesto y Anderson sinonimizaron con Reptilia, ya que este último es más conocido y más utilizado. Sin embargo, a diferencia de la mayoría de las definiciones anteriores de Reptilia, la definición de Modesto y Anderson incluye aves, ya que están dentro del clado que incluye tanto a los lagartos como a los cocodrilos.
Taxonomía
Clasificación para ordenar el nivel de los reptiles, después de Benton, 2014.
- Clase Reptilia
- † Subclase Parareptilia
- † Ordenar Pareiasauromorpha
- Subclase Eureptilia
- Infraclass Diapsida
- † Orden Younginiformes
- Infraclass Neodiapsida
- Ordenar Testudinata (tortugas)
- Infraclass Lepidosauromorpha
- Infrasubclass Sin nombre
- † Infraclass Ichthyosauria
- † Orden Thalattosauria
- Superorden Lepidosauriformes
- Orden Rhynchocephalia (tuatara)
- Ordenar Squamata (lagartos y serpientes)
- † Infrasubclass Sauropterygia
- † Orden Placodoncia
- † Orden Eosauropterygia
- † Orden Plesiosauria
- Infrasubclass Sin nombre
- Infraclass Archosauromorpha
- † Orden Rhynchosauria
- † Orden Protorosauria
- † Orden Phytosauria
- División Archosauriformes
- Subdivisión Archosauria
- Superorden Crocodylomorpha
- Orden Crocodilia
- Infradivision Avemetatarsalia
- Infrasubdivision Ornithodira
- † Orden Pterosauria
- Superorden Dinosauria
- Orden Saurischia (incluido Clade Aves )
- † Orden Ornithischia
- Infrasubdivision Ornithodira
- Superorden Crocodylomorpha
- Subdivisión Archosauria
- Infraclass Diapsida
- † Subclase Parareptilia
Filogenia
El cladograma presentado aquí ilustra el "árbol genealógico" de los reptiles, y sigue una versión simplificada de las relaciones encontradas por MS Lee, en 2013. Todos los estudios genéticos han respaldado la hipótesis de que las tortugas son diapsidas; algunos han colocado tortugas dentro de los archosauriformes, aunque algunos han recuperado las tortugas como lepidosauriformes en su lugar. El cladograma a continuación utilizó una combinación de datos genéticos (moleculares) y fósiles (morfológicos) para obtener sus resultados.
| Amniota |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||







La posición de las tortugas
La ubicación de las tortugas ha sido históricamente muy variable. Clásicamente, se consideraba que las tortugas estaban relacionadas con reptiles anapsid primitivos. El trabajo molecular generalmente ha colocado tortugas dentro de los diápsidos. Hasta ahora, tres genomas de tortugas han sido secuenciados. Los resultados ubican a las tortugas como un clado hermano de los arcosaurios, el grupo que incluye cocodrilos, dinosaurios y aves.
Historia Evolutiva
Origen de los reptiles


El origen de los reptiles se encuentra alrededor de 310-320 millones de años atrás, en los humeantes pantanos de finales del período Carbonífero, cuando los primeros reptiles evolucionaron a partir de reptilomorfos avanzados.
El animal más antiguo conocido que pudo haber sido un amniote es Casineria (aunque puede haber sido un temnospondilo). Una serie de huellas de los estratos fósiles de Nueva Escocia fechadas en 315 Ma muestran dedos de reptil típicos e impresiones de escamas. Estas huellas se atribuyen a Hylonomus , el reptil indiscutible más antiguo que se conoce. Era un animal pequeño, parecido a una lagartija, de unos 20 a 30 centímetros (7.9 a 11.8 pulgadas) de largo, con numerosos dientes afilados que indicaban una dieta insectívora. Otros ejemplos incluyen Westlothiana (por el momento considerado un reptiliomorph en lugar de un verdadero amniote) y Paleothyris , ambos de construcción similar y hábito probablemente similar.
Alza de los reptiles
Los primeros amniotas, incluidos los reptiles de tallo (los amniotas más cercanos a los reptiles modernos que a los mamíferos), fueron en gran parte eclipsados por los tetrápodos de tallo más grandes, como Cochleosaurus, y se mantuvo como una pequeña y discreta parte de la fauna hasta el colapso del bosque lluvioso carbonífero. Este colapso repentino afectó a varios grupos grandes. Los tetrápodos primitivos fueron particularmente devastados, mientras que los reptiles de tallo se comportaron mejor, estando ecológicamente adaptados a las condiciones más secas que siguieron. Los tetrápodos primitivos, como los anfibios modernos, necesitan regresar al agua para desovar; en cambio, los amniotes, como los reptiles modernos, cuyos huevos poseen un caparazón que les permite colocarse en tierra, se adaptaron mejor a las nuevas condiciones. Amniotes adquirió nuevos nichos a un ritmo más rápido que antes del colapso y a un ritmo mucho más rápido que los tetrápodos primitivos. Adquirieron nuevas estrategias de alimentación que incluyen herbivoría y carnivoría, anteriormente solo habían sido insectívoros y piscívoros. Desde este punto en adelante, los reptiles dominaban las comunidades y tenían una mayor diversidad que los tetrápodos primitivos, preparando el escenario para el Mesozoico (conocido como la Era de los Reptiles). Uno de los reptiles primarios más conocidos es Mesosaurus, un género del Pérmico temprano que había regresado al agua, alimentándose de peces.
Anapsidos, sinapsidos, diapsidos y sauropsidos

Tradicionalmente se suponía que los primeros reptiles conservaban un cráneo anápsido heredado de sus antepasados. Este tipo de cráneo tiene un techo de calavera con solo orificios para las fosas nasales, los ojos y un ojo pineal. Los descubrimientos de aberturas similares a sinapsidos (ver más abajo) en el techo del cráneo de los cráneos de varios miembros de Parareptilia (el clado que contiene la mayoría de los amniotas llamados tradicionalmente "anapsidos"), incluyendo lanthanosuchoides, millerettids, bolosaurids, algunos nyterooleterids, algunos procolofonoides y al menos algunos mesosaurios lo hicieron más ambiguo y actualmente no está claro si el amniote ancestral tenía un cráneo similar a la anápsida o sinápsido. Estos animales se conocen tradicionalmente como "anápsidos" y forman una reserva básica parafilética a partir de la cual evolucionaron otros grupos. Muy poco después de que aparecieron los primeros amniotas, un linaje llamado Synapsida se separó; este grupo se caracterizó por una abertura temporal en el cráneo detrás de cada ojo para dar lugar al movimiento del músculo de la mandíbula. Estos son los "amniotas similares a los mamíferos", o mamíferos de tallo, que más tarde dieron origen a los verdaderos mamíferos. Poco después, otro grupo desarrolló un rasgo similar, esta vez con una doble apertura detrás de cada ojo, ganándose el nombre Diapsida ("dos arcos"). La función de los agujeros en estos grupos era aligerar el cráneo y dejar espacio para que los músculos de la mandíbula se muevan, lo que permite una mordida más potente. que más tarde dio origen a los verdaderos mamíferos. Poco después, otro grupo desarrolló un rasgo similar, esta vez con una doble apertura detrás de cada ojo, ganándose el nombre Diapsida ("dos arcos"). La función de los agujeros en estos grupos era aligerar el cráneo y dejar espacio para que los músculos de la mandíbula se muevan, lo que permite una mordida más potente. que más tarde dio origen a los verdaderos mamíferos. Poco después, otro grupo desarrolló un rasgo similar, esta vez con una doble apertura detrás de cada ojo, ganándose el nombre Diapsida ("dos arcos"). La función de los agujeros en estos grupos era aligerar el cráneo y dejar espacio para que los músculos de la mandíbula se muevan, lo que permite una mordida más potente.
Tradicionalmente, se creía que las tortugas sobrevivían parareptiles, sobre la base de su estructura de cráneo anápsida, que se suponía era un rasgo primitivo. La razón de esta clasificación ha sido discutida, y algunos argumentan que las tortugas son diapsidos que desarrollaron cráneos de anápsidos para mejorar su armadura. Los estudios filogenéticos morfológicos posteriores con esto en mente colocaron a las tortugas firmemente dentro de Diapsida. Todos los estudios moleculares han confirmado la ubicación de las tortugas dentro de los diápsidos, más comúnmente como un grupo hermano de los archosaurios existentes.
Reptiles pérmicos
Con el cierre del Carbonífero, los amniotes se convirtieron en la fauna tetrápoda dominante. Mientras todavía existían reptiliomorfos terrestres primitivos, los amniotes sinápsidos desarrollaron la primera megafauna verdaderamente terrestre (animales gigantes) en forma de pelicosaurios, como Edaphosaurus y el Dimetrodon carnívoro . A mediados del período Pérmico, el clima se volvió más seco, lo que provocó un cambio en la fauna: los pelicosaurios fueron reemplazados por los terápsidos.
Los parareptiles, cuyos enormes techos de calavera no tenían agujeros postorbitales, continuaron y florecieron en todo el Pérmico. Los parareptiles pareiasaurianos alcanzaron proporciones gigantes en el Pérmico tardío, y eventualmente desaparecieron al final del período (las tortugas son posibles sobrevivientes).
Temprano en el período, los reptiles modernos, o reptiles del grupo corona, evolucionaron y se dividieron en dos linajes principales: los Archosauromorpha (antepasados de tortugas, cocodrilos y dinosaurios) y los Lepidosauromorpha (predecesores de los lagartos y tuataras modernos). Ambos grupos permanecieron como lagartos y relativamente pequeños y discretos durante el Pérmico.
Reptiles mesozoicos
El cierre del Pérmico vio la mayor extinción masiva conocida (véase el evento de extinción Pérmico-Triásico), un evento prolongado por la combinación de dos o más pulsos de extinción distintos. La mayor parte de la megafauna parapetáptica y sinápsida anterior desapareció, siendo reemplazada por los verdaderos reptiles, particularmente los archosauromorfos. Estos se caracterizaron por patas traseras alargadas y una pose erecta, las primeras formas parecían algo así como cocodrilos de largas patas. Los arcosaurios se convirtieron en el grupo dominante durante el período Triásico, aunque transcurrieron 30 millones de años antes de que su diversidad fuera tan grande como la de los animales que vivían en el Pérmico. Los Archosaurios se convirtieron en los famosos dinosaurios y pterosaurios, así como en los ancestros de los cocodrilos. Dado que los reptiles, los primeros rauisuquios y luego los dinosaurios, dominaron la era mesozoica, el intervalo se conoce popularmente como la "Era de los reptiles". Los dinosaurios también desarrollaron formas más pequeñas, incluyendo los terópodos más pequeños que llevan plumas. En el período Cretácico, estos dieron lugar a las primeras aves verdaderas.
El grupo hermano de Archosauromorpha es Lepidosauromorpha, que contiene lagartos y tuataras, así como sus parientes fósiles. Lepidosauromorpha contenía al menos un grupo principal de reptiles marinos mesozoicos: los mosasauros, que vivieron durante el período Cretácico. La ubicación filogenética de otros grupos principales de reptiles marinos fósiles, los ictiópteros (incluidos ictiosaurios) y los sauropterigios, que se desarrollaron a comienzos del Triásico, es más controvertida. Diferentes autores relacionaron estos grupos con lepidosauromorfos o con archosauromorfos, y también se argumentó que los ictiopérgicos eran diapsidos que no pertenecían al clado menos inclusivo que contenía lepidosauromorfos y archosauromorfos.
Reptiles cenozoicos

El cierre del período Cretácico vio la desaparición de la megafauna reptiliana de la era Mesozoica (ver el evento de extinción Cretácico-Paleógeno). De los grandes reptiles marinos, solo quedaron las tortugas marinas; y de los grandes reptiles no marinos, solo los cocodrilos semiacuáticos y los coristoderos ampliamente similares sobrevivieron a la extinción, y estos últimos se extinguieron en el Mioceno. De la gran cantidad de dinosaurios que dominaron el Mesozoico, solo sobrevivieron las pequeñas aves de pico. Este dramático patrón de extinción al final del Mesozoico condujo al Cenozoico. Mamíferos y pájaros llenaron los vacíos nichos dejados por la megafauna reptiliana y, mientras la diversificación de los reptiles disminuía, la diversificación de aves y mamíferos tomó un giro exponencial. Sin embargo, los reptiles todavía eran componentes importantes de la megafauna, particularmente en forma de tortugas gigantes.
Después de la extinción de la mayoría de las líneas de reptiles marinos y de archosaurios a fines del Cretácico, la diversificación de reptiles continuó durante todo el Cenozoico. Los Squamates recibieron un golpe masivo durante el evento KT, recuperándose solo diez millones de años después, pero experimentaron un gran evento de radiación una vez que se recuperaron, y hoy los escuamatos constituyen la mayoría de los reptiles vivos (> 95%). Se conocen aproximadamente 10.000 especies de reptiles tradicionales existentes, y las aves agregan aproximadamente 10.000 más, casi el doble de mamíferos, representados por unas 5.700 especies vivas (excluidas las especies domesticadas).
Morfología y fisiología
Circulación

Todos los escuamatos y tortugas tienen un corazón de tres cámaras que consta de dos aurículas, un ventrículo dividido de forma variable y dos aortas que conducen a la circulación sistémica. El grado de mezcla de sangre oxigenada y desoxigenada en el corazón de tres cámaras varía según la especie y el estado fisiológico. En diferentes condiciones, la sangre desoxigenada se puede derivar al cuerpo o la sangre oxigenada se puede derivar a los pulmones. Esta variación en el flujo sanguíneo se ha hipotetizado para permitir una termorregulación más efectiva y tiempos de inmersión más largos para las especies acuáticas, pero no se ha demostrado que sea una ventaja de aptitud física.
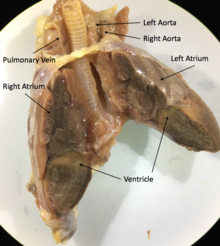
Por ejemplo, los corazones de Iguana, como la mayoría de los corazones de los escuamatos, se componen de tres cámaras con dos aorta y un ventrículo, músculos cardiacos involuntarios. Las principales estructuras del corazón son el seno venoso, el marcapasos, la aurícula izquierda, el atruim derecho, la válvula auriculoventricular, la cavidad venosa, el cavum arterioso, el cavum pulmonar, la cresta muscular, la cresta ventricular, las venas pulmonares y los pares arcos aórticos.
Algunas especies de squamate (por ejemplo, pitones y lagartos monitores) tienen corazones de tres cámaras que se convierten funcionalmente en corazones de cuatro cámaras durante la contracción. Esto es posible gracias a un reborde muscular que subdivide el ventrículo durante la diástole ventricular y lo divide por completo durante la sístole ventricular. Debido a esta cresta, algunos de estos escuamatos son capaces de producir diferenciales de presión ventricular que son equivalentes a los observados en corazones de mamíferos y aves.
Los cocodrilos tienen un corazón anatómicamente de cuatro cámaras, similar a las aves, pero también tienen dos aortas sistémicas y, por lo tanto, son capaces de pasar por alto su circulación pulmonar.
Metabolismo
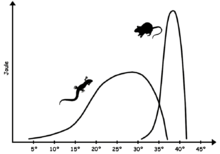
Los reptiles no aviares modernos exhiben alguna forma de sangre fría (es decir, una mezcla de poiquilotermia, ectotermia y bradymetabolism) por lo que tienen medios fisiológicos limitados para mantener la temperatura corporal constante y, a menudo dependen de fuentes externas de calor. Debido a una temperatura central menos estable que las aves y los mamíferos, la bioquímica de los reptiles requiere enzimas capaces de mantener la eficiencia en un rango mayor de temperaturas que en el caso de los animales de sangre caliente. El rango óptimo de temperatura corporal varía con las especies, pero típicamente es inferior al de los animales de sangre caliente; para muchos lagartos, cae en el rango de 24 ° -35 ° C (75 ° -95 ° F), mientras que especies extremas adaptadas al calor, como la iguana del desierto estadounidense Dipsosaurus dorsalis, puede tener temperaturas fisiológicas óptimas en el rango de mamíferos, entre 35 ° y 40 ° C (95 ° y 104 ° F). Si bien la temperatura óptima se encuentra con frecuencia cuando el animal está activo, el metabolismo basal bajo hace que la temperatura corporal baje rápidamente cuando el animal está inactivo.
Como en todos los animales, la acción muscular reptil produce calor. En grandes reptiles, como las tortugas laúd, la baja relación superficie-volumen permite que este calor producido metabólicamente mantenga a los animales más calientes que su entorno a pesar de que no tienen un metabolismo de sangre caliente. Esta forma de homeotermia se llama gigantothermy; se ha sugerido que era común en dinosaurios grandes y otros reptiles extintos de cuerpo grande.
El beneficio de un bajo metabolismo en reposo es que requiere mucho menos combustible para mantener las funciones corporales. Al usar variaciones de temperatura en su entorno, o al permanecer fríos cuando no necesitan moverse, los reptiles pueden ahorrar una cantidad considerable de energía en comparación con los animales endotérmicos del mismo tamaño. Un cocodrilo necesita entre un décimo y un quinto de la comida necesaria para un león del mismo peso y puede vivir medio año sin comer. Los menores requerimientos de alimentos y el metabolismo adaptativo permiten a los reptiles dominar la vida animal en regiones donde la disponibilidad neta de calorías es demasiado baja para mantener a los mamíferos y aves de gran tamaño.
En general, se supone que los reptiles no pueden producir la producción sostenida de alta energía necesaria para las persecuciones de larga distancia o el vuelo. Una mayor capacidad energética podría haber sido responsable de la evolución de la sangre caliente en aves y mamíferos. Sin embargo, la investigación de las correlaciones entre la capacidad activa y la termofisiología muestra una relación débil. La mayoría de los reptiles existentes son carnívoros con una estrategia de alimentación de sentarse y esperar; si los reptiles son de sangre fría debido a su ecología no está claro. Los estudios energéticos en algunos reptiles han mostrado capacidades activas iguales o mayores que los animales de sangre caliente de tamaño similar.
Sistema respiratorio
Todos los reptiles respiran usando los pulmones. Las tortugas acuáticas han desarrollado una piel más permeable, y algunas especies han modificado su cloaca para aumentar el área de intercambio de gases. Incluso con estas adaptaciones, la respiración nunca se logra completamente sin los pulmones. La ventilación pulmonar se realiza de forma diferente en cada grupo principal de reptiles. En los escuamatos, los pulmones están ventilados casi exclusivamente por la musculatura axial. Esta es también la misma musculatura que se usa durante la locomoción. Debido a esta restricción, la mayoría de los escuamatos se ven obligados a contener la respiración durante carreras intensas. Algunos, sin embargo, han encontrado una forma de evitarlo. Varanids, y algunas otras especies del lagarto, emplean el bombeo bucal como complemento a su "respiración axial normal". Esto permite que los animales llenen completamente sus pulmones durante una locomoción intensa, y así permanecen aeróbicamente activos por un largo tiempo. Se sabe que los lagartos Tegu poseen un proto-diafragma, que separa la cavidad pulmonar de la cavidad visceral. Si bien no es capaz de moverse, sí permite una mayor inflación pulmonar, al quitar el peso de las vísceras de los pulmones.
Los cocodrilos realmente tienen un diafragma muscular que es análogo al diafragma de los mamíferos. La diferencia es que los músculos del diafragma de cocodrilo empujan hacia atrás el pubis (parte de la pelvis, que es movible en los cocodrilos), lo que hace que el hígado baje, liberando así espacio para que los pulmones se expandan. Este tipo de configuración diafragmática se conoce como el "pistón hepático". Las vías respiratorias forman una serie de cámaras tubulares dobles dentro de cada pulmón. Al inhalar y exhalar, el aire se mueve a través de las vías respiratorias en la misma dirección, creando así un flujo de aire unidireccional a través de los pulmones. Un sistema similar se encuentra en aves, lagartos monitores e iguanas.
La mayoría de los reptiles carecen de un paladar secundario, lo que significa que deben contener la respiración mientras tragan. Los cocodrílidos han desarrollado un paladar óseo secundario que les permite seguir respirando mientras permanecen sumergidos (y proteger sus cerebros contra el daño de las presas que luchan). Eslizones (familia Scincidae) también han desarrollado un paladar óseo secundario, en diversos grados. Las serpientes tomaron un enfoque diferente y extendieron su tráquea en su lugar. Su extensión traqueal sobresale como una paja carnosa, y les permite a estos animales tragar presas grandes sin sufrir de asfixia.
Tortugas y tortugas

Cómo respiran tortugas y tortugas ha sido objeto de numerosos estudios. Hasta la fecha, solo unas pocas especies se han estudiado lo suficiente como para tener una idea de cómo respiran esas tortugas. Los resultados variados indican que las tortugas y las tortugas han encontrado una variedad de soluciones a este problema.
La dificultad es que la mayoría de las conchas de tortuga son rígidas y no permiten el tipo de expansión y contracción que otros amniotas usan para ventilar sus pulmones. Algunas tortugas, como el aleta india ( Lissemys punctata), tiene una hoja de músculo que envuelve los pulmones. Cuando se contrae, la tortuga puede exhalar. Cuando está en reposo, la tortuga puede retraer las extremidades hacia la cavidad corporal y expulsar el aire de los pulmones. Cuando la tortuga prolonga sus extremidades, la presión dentro de los pulmones se reduce y la tortuga puede aspirar aire. Los pulmones de tortuga se unen al interior de la parte superior de la caparazón (caparazón), con la parte inferior de los pulmones unidas (a través de tejido) al resto de las vísceras. Al usar una serie de músculos especiales (más o menos equivalente a un diafragma), las tortugas son capaces de empujar sus vísceras hacia arriba y hacia abajo, lo que resulta en una respiración efectiva, ya que muchos de estos músculos tienen puntos de unión en conjunción con sus extremidades anteriores (de hecho, muchas los músculos se expanden en los bolsillos de las extremidades durante la contracción).
La respiración durante la locomoción se ha estudiado en tres especies, y muestran diferentes patrones. Las tortugas marinas verdes adultas no respiran mientras se mueven a lo largo de sus playas de anidación. Mantienen la respiración durante la locomoción terrestre y respiran en los combates mientras descansan. Las tortugas de caja norteamericanas respiran continuamente durante la locomoción, y el ciclo de ventilación no está coordinado con los movimientos de las extremidades. Esto se debe a que usan sus músculos abdominales para respirar durante la locomoción. La última especie que se ha estudiado es el deslizador de orejas rojas, que también respira durante la locomoción, pero respira durante la locomoción más leve que durante las pausas pequeñas entre combates locomotores, lo que indica que puede haber interferencia mecánica entre los movimientos de las extremidades y el aparato de respiración.
Piel

La piel reptil está cubierta por una epidermis córnea, lo que la hace estanca y permite que los reptiles vivan en tierra firme, a diferencia de los anfibios. En comparación con la piel de mamíferos, la de los reptiles es bastante delgada y carece de la gruesa capa dérmica que produce el cuero en los mamíferos. Las partes expuestas de los reptiles están protegidas por escamas o escudos, a veces con una base ósea, formando una armadura. En lepidosaurios, como lagartos y serpientes, toda la piel está cubierta por escamas epidérmicas superpuestas. Se creía que tales escalas eran típicas de la clase Reptilia en su conjunto, pero ahora se sabe que ocurren solo en lepidosaurios. Las escamas encontradas en las tortugas y cocodrilos son de origen dérmico, en lugar de epidérmico, y se denominan apropiadamente escudos. En las tortugas, el cuerpo está escondido dentro de una caparazón duro compuesto de escudetes fusionados.
Al carecer de una dermis espesa, el cuero de reptil no es tan fuerte como el cuero de los mamíferos. Se utiliza en artículos de cuero con fines decorativos para zapatos, cinturones y bolsos, especialmente piel de cocodrilo.
Derramamiento. Los reptiles mudan su piel a través de un proceso llamado ecdisis que ocurre continuamente a lo largo de su vida. En particular, los reptiles más jóvenes tienden a arrojar una vez cada 5-6 semanas mientras que los adultos arrojan 3-4 veces al año. Los reptiles más jóvenes arrojan más debido a su rápida tasa de crecimiento. Una vez que el tamaño completo, la frecuencia de desprendimiento disminuye drásticamente. El proceso de ecdisis implica formar una nueva capa de piel debajo de la anterior. Las enzimas proteolíticas y el líquido linfático se secretan entre las capas de piel nuevas y viejas. En consecuencia, esto levanta la piel vieja de la nueva permitiendo que ocurra el desprendimiento. Las serpientes se arrojarán de la cabeza a la cola mientras que las lagartijas se mudarán en un "patrón irregular". Dysecdysis, una enfermedad común de la piel en serpientes y lagartos, ocurrirá cuando falla la ecdisis o derramamiento. Existen numerosas razones por las que el desprendimiento falla y pueden estar relacionados con una humedad y temperatura inadecuadas, deficiencias nutricionales, deshidratación y lesiones traumáticas. Las deficiencias nutricionales disminuyen las enzimas proteolíticas, mientras que la deshidratación reduce los fluidos linfáticos para separar las capas de la piel. Las lesiones traumáticas, por otro lado, forman cicatrices que no permitirán que se formen nuevas escalas e interrumpan el proceso de ecdisis.
Excreción
La excreción se realiza principalmente por dos riñones pequeños. En los diápsidos, el ácido úrico es el principal producto de desecho nitrogenado; las tortugas, como los mamíferos, excretan principalmente urea. A diferencia de los riñones de mamíferos y aves, los riñones de reptiles no pueden producir orina líquida más concentrada que sus fluidos corporales. Esto se debe a que carecen de una estructura especializada llamada asa de Henle, que está presente en las nefronas de aves y mamíferos. Debido a esto, muchos reptiles usan el colon para ayudar en la reabsorción de agua. Algunos también pueden absorber el agua almacenada en la vejiga. El exceso de sales también se excreta por las glándulas salinas nasales y linguales en algunos reptiles.
En todos los reptiles, los conductos urinogenitales y el ano se vacían en un órgano llamado cloaca. En algunos reptiles, una pared medioventral en la cloaca puede abrirse a una vejiga urinaria, pero no a todos. Está presente en todas las tortugas y tortugas, así como en la mayoría de los lagartos, pero falta en el lagarto monitor, los lagartos sin patas. Está ausente en las serpientes, los caimanes y los cocodrilos.
Muchas tortugas, tortugas y lagartos tienen vejigas proporcionalmente muy grandes. Charles Darwin notó que la tortuga de Galápagos tenía una vejiga que podía almacenar hasta un 20% de su peso corporal. Tales adaptaciones son el resultado de entornos tales como islas remotas y desiertos donde el agua es muy escasa. Otros reptiles que habitan en el desierto tienen grandes vejigas que pueden almacenar un reservorio de agua a largo plazo durante varios meses y ayudan a la osmorregulación.
Las tortugas tienen dos o más vejigas urinarias accesorias, localizadas lateralmente al cuello de la vejiga urinaria y dorsal al pubis, que ocupan una parte importante de la cavidad corporal. Su vejiga también suele ser bilobulada con una sección izquierda y derecha. La sección derecha se encuentra debajo del hígado, lo que evita que las piedras grandes permanezcan en ese lado, mientras que la sección izquierda es más propensa a tener cálculos.
Digestión

La mayoría de los reptiles son insectívoros o carnívoros y tienen tractos digestivos simples y comparativamente cortos debido a que la carne es bastante simple de descomponer y digerir. La digestión es más lenta que en los mamíferos, lo que refleja su menor metabolismo en reposo y su incapacidad para dividir y masticar sus alimentos. Su metabolismo poiquilotermo tiene requisitos de energía muy bajos, lo que permite que los reptiles grandes, como los cocodrilos y los grandes constrictores, puedan vivir de una sola gran comida durante meses, digiriéndola lentamente.
Mientras que los reptiles modernos son predominantemente carnívoros, durante la historia temprana de los reptiles varios grupos produjeron alguna megafauna herbívora: en el Paleozoico, los pareiasaurios; y en el Mesozoico varias líneas de dinosaurios. Hoy en día, las tortugas son el único grupo de reptiles predominantemente herbívoros, pero varias líneas de agamas e iguanas han evolucionado para vivir total o parcialmente en las plantas.
Los reptiles herbívoros enfrentan los mismos problemas de masticación que los mamíferos herbívoros pero, al carecer de los dientes complejos de los mamíferos, muchas especies tragan rocas y guijarros (llamados gastrolitos) para ayudar en la digestión: las rocas se lavan en el estómago, ayudando a moler la planta importar. Los gastrolitos fósiles se han encontrado asociados tanto con ornitópodos como con saurópodos, aunque se discute si realmente funcionaron como una fábrica gástrica en este último. Los cocodrilos de agua salada también usan gastrolitos como lastre, estabilizándolos en el agua o ayudándolos a bucear. Se ha sugerido una doble función como estabilizador de lastre y digestión para gastrolitos encontrados en plesiosaurios.
Nervios
El sistema nervioso reptil contiene la misma parte básica del cerebro anfibio, pero el cerebro de reptil y el cerebelo son un poco más grandes. La mayoría de los órganos de los sentidos típicos están bien desarrollados, con ciertas excepciones, sobre todo la falta de orejas externas de la serpiente (los oídos medios e internos están presentes). Hay doce pares de nervios craneales. Debido a su cóclea corta, los reptiles usan la afinación eléctrica para expandir su rango de frecuencias audibles.
Inteligencia
Los reptiles generalmente se consideran menos inteligentes que los mamíferos y las aves. El tamaño de su cerebro en relación con su cuerpo es mucho menor que el de los mamíferos, el cociente de encefalización es aproximadamente una décima parte del de los mamíferos, aunque los reptiles más grandes pueden mostrar un desarrollo cerebral más complejo. Los lagartos más grandes, como los monitores, son conocidos por exhibir un comportamiento complejo, incluida la cooperación. Los cocodrilos tienen cerebros relativamente más grandes y muestran una estructura social bastante compleja. Incluso se sabe que el dragón de Komodo participa en el juego, al igual que las tortugas, que también se consideran criaturas sociales, y algunas veces cambian de monogamia a promiscuidad en su comportamiento sexual. Un estudio encontró que las tortugas de madera eran mejores que las ratas blancas para aprender a navegar en laberintos.
Visión
La mayoría de los reptiles son animales diurnos. La visión se adapta típicamente a las condiciones de luz diurna, con visión de color y percepción de profundidad visual más avanzada que en anfibios y en la mayoría de los mamíferos. En algunas especies, como las serpientes ciegas, la visión se reduce.
Algunas serpientes tienen conjuntos adicionales de órganos visuales (en el sentido más amplio de la palabra) en forma de fosas sensibles a la radiación infrarroja (calor). Tales pozos sensibles al calor están particularmente bien desarrollados en las víboras, pero también se encuentran en boas y pitones. Estas fosas permiten a las serpientes detectar el calor corporal de aves y mamíferos, lo que permite a las víboras de caza cazar roedores en la oscuridad.
Reproducción




Los reptiles generalmente se reproducen sexualmente, aunque algunos son capaces de reproducción asexual. Toda la actividad reproductiva ocurre a través de la cloaca, la única salida / entrada en la base de la cola donde también se eliminan los desechos. La mayoría de los reptiles tienen órganos copulatorios, que generalmente se retraen o se invierten y se almacenan dentro del cuerpo. En las tortugas y los cocodrilos, el macho tiene un único pene mediano, mientras que los escamosos, incluidas las serpientes y los lagartos, poseen un par de hemipenes, de los cuales solo uno se usa normalmente en cada sesión. Los tuatara, sin embargo, carecen de órganos copulatorios, por lo que el macho y la hembra simplemente presionan sus cloacas juntas a medida que el macho descarga el esperma.
La mayoría de los reptiles ponen huevos amnióticos cubiertos con conchas correosas o calcáreas. Un amnios, corion y alantoides están presentes durante la vida embrionaria. La cáscara de huevo (1) protege el embrión de cocodrilo (11) y evita que se seque, pero es flexible para permitir el intercambio de gases. El corion (6) ayuda al intercambio de gases entre el interior y el exterior del huevo. Permite que el dióxido de carbono salga del huevo y del oxígeno gaseoso para ingresar al huevo. La albúmina (9) protege aún más el embrión y sirve como un depósito de agua y proteínas. El alantoides (8) es un saco que recoge los desechos metabólicos producidos por el embrión. El saco amniótico (10) contiene líquido amniótico (12) que protege y amortigua el embrión. El amnios (5) ayuda a la osmorregulación y sirve como depósito de agua salada. El saco vitelino (2) que rodea la yema (3) contiene proteínas y nutrientes ricos en grasa que son absorbidos por el embrión a través de los vasos (4) que permiten que el embrión crezca y se metabolice. El espacio de aire (7) proporciona al embrión oxígeno mientras está incubando. Esto asegura que el embrión no se sofocará mientras eclosiona. No hay etapas larvales de desarrollo. Viviparity y ovoviviparity han evolucionado en muchos clados extintos de reptiles y en squamates. En este último grupo, muchas especies, incluidas todas las boas y la mayoría de las víboras, utilizan este modo de reproducción. El grado de viviparidad varía; algunas especies simplemente retienen los huevos hasta justo antes de la eclosión, otras proporcionan alimento materno para complementar la yema y otras carecen de yema y proporcionan todos los nutrientes a través de una estructura similar a la placenta de los mamíferos. El primer caso documentado de viviparidad en reptiles es el de los mesosaurios del Pérmico temprano, aunque algunos individuos o taxones en ese clado también pueden haber sido ovíparos debido a que también se ha encontrado un supuesto huevo aislado. Varios grupos de reptiles marinos mesozoicos también exhibieron viviparidad, como mosasaurios, ictiosauros y Sauropterygia, un grupo que incluye pachypleurosaurs y Plesiosauria.
La reproducción asexual ha sido identificada en escamosos en seis familias de lagartos y una serpiente. En algunas especies de escuamatos, una población de hembras puede producir un clon diploide unisexual de la madre. Esta forma de reproducción asexual, llamada partenogénesis, se produce en varias especies de gecko, y está especialmente extendida en los teiidos (especialmente Aspidocelis ) y los lacécidos ( Lacerta ). En cautiverio, los dragones de Komodo (Varanidae) se han reproducido por partenogénesis.
Se sospecha que existen especies partenogenéticas entre camaleones, ágamas, xantusiidos y tiflopidos.
Algunos reptiles exhiben la determinación del sexo dependiente de la temperatura (TDSD), en el cual la temperatura de incubación determina si un huevo en particular eclosiona como macho o hembra. TDSD es más común en tortugas y cocodrilos, pero también ocurre en lagartos y tuatara. Hasta la fecha, no ha habido confirmación de si TDSD ocurre en serpientes.
Mecanismos de defensa
Muchos pequeños reptiles, como las serpientes y los lagartos que viven en el suelo o en el agua, son vulnerables a ser depredados por todo tipo de animales carnívoros. Por lo tanto, evitar es la forma más común de defensa en reptiles. A la primera señal de peligro, la mayoría de las serpientes y lagartos se arrastran hacia la maleza, y las tortugas y los cocodrilos se sumergirán en el agua y se perderán de vista.
Camuflaje y advertencia

Los reptiles tienden a evitar la confrontación a través del camuflaje. Dos grupos principales de depredadores de reptiles son aves y otros reptiles, ambos con una visión del color bien desarrollada. Por lo tanto, las pieles de muchos reptiles tienen una coloración críptica de gris liso o moteado, verde y marrón que les permite fundirse en el fondo de su entorno natural. Ayudado por la capacidad de los reptiles para permanecer inmóviles durante largos períodos de tiempo, el camuflaje de muchas serpientes es tan eficaz que las personas o los animales domésticos son más comúnmente mordidos porque accidentalmente los pisa.
Cuando el camuflaje no los protege, los eslizones de lengua azul intentarán protegerse de los atacantes mostrando sus lenguas azules, y el lagarto de cuello volante exhibirá su volante de colores brillantes. Estas mismas pantallas se usan en disputas territoriales y durante el cortejo. Si el peligro surge tan repentinamente que el vuelo es inútil, cocodrilos, tortugas, algunas lagartijas y algunas serpientes silban ruidosamente cuando se enfrentan con un enemigo. Las serpientes de cascabel hacen vibrar rápidamente la punta de la cola, que está compuesta por una serie de cuentas huecas anidadas para protegerse del peligro que se avecina.
En contraste con la coloración gris normal de la mayoría de los reptiles, las lagartijas del género Heloderma (el monstruo de Gila y la lagartija de cuentas) y muchas de las serpientes de coral tienen una coloración de advertencia de alto contraste, advirtiendo a los posibles depredadores que son venenosas. Las especies de serpientes venenosas de América del Norte tienen marcas coloridas similares a las de la serpiente de coral, un ejemplo frecuentemente citado de mímica batesiana.
Defensa alternativa en serpientes
El camuflaje no siempre engaña a un depredador. Cuando son atrapados, las especies de serpientes adoptan diferentes tácticas defensivas y usan un conjunto complicado de comportamientos cuando son atacados. Algunos primero elevan su cabeza y extienden la piel de su cuello en un esfuerzo por verse grandes y amenazantes. El fracaso de esta estrategia puede conducir a otras medidas practicadas particularmente por cobras, víboras y especies estrechamente relacionadas, que usan veneno para atacar. El veneno es saliva modificada, liberada a través de colmillos de una glándula venenosa. Algunas serpientes no venenosas, como las serpientes hognose americanas o las serpientes de hierba europeas, mueren cuando están en peligro; algunos, incluida la serpiente de hierba, exudan un líquido maloliente para disuadir a los atacantes.
Defensa en cocodrilos
Cuando un cocodrílido está preocupado por su seguridad, se abrirá para exponer los dientes y la lengua amarilla. Si esto no funciona, el cocodrílido se pone un poco más agitado y por lo general comienza a hacer silbidos. Después de esto, el cocodrilo comenzará a cambiar dramáticamente su postura para parecer más intimidante. El cuerpo está inflado para aumentar el tamaño aparente. Si es absolutamente necesario, puede decidir atacar a un enemigo.

Algunas especies intentan morder de inmediato. Algunos utilizarán sus cabezas como mazos y literalmente aplastarán a un oponente, algunos se apresurarán o nadarán hacia la amenaza desde la distancia, incluso persiguiendo al oponente hacia la tierra o galopando tras él. El arma principal en todos los cocodrilos es la picadura, que puede generar una fuerza de mordida muy alta. Muchas especies también poseen dientes similares a caninos. Estos se usan principalmente para capturar presas, pero también se usan para combatir y exhibir.
Derramando y regenerando colas
Los gecos, los eslizones y otras lagartijas que son capturadas por la cola perderán parte de la estructura de la cola a través de un proceso llamado autotomía y así podrán huir. La cola separada continuará moviéndose, creando una sensación engañosa de lucha continua y distrayendo la atención del depredador del animal que está huyendo. Las colas desprendidas de geckos leopardo se pueden mover hasta por 20 minutos. En muchas especies, las colas son de un color separado y dramáticamente más intenso que el resto del cuerpo para animar a los depredadores potenciales a golpear primero la cola. En el eslizón de guijarros y en algunas especies de gecos, la cola es corta y ancha y se asemeja a la cabeza, por lo que los depredadores pueden atacarla en lugar de la parte delantera más vulnerable.
Los reptiles que son capaces de arrojar sus colas pueden regenerarlos parcialmente durante un período de semanas. Sin embargo, la nueva sección contendrá cartílago en lugar de hueso, y nunca crecerá a la misma longitud que la cola original. A menudo también se decolora claramente en comparación con el resto del cuerpo y puede carecer de algunas de las características de esculpir externas vistas en la cola original.
Relaciones con los humanos
En culturas y religiones

Los dinosaurios han sido ampliamente representados en la cultura desde que el paleontólogo inglés Richard Owen acuñó el nombre de dinosaurio en 1842. Ya en 1854, los dinosaurios Crystal Palace estaban en exhibición para el público en el sur de Londres. Un dinosaurio apareció en la literatura incluso antes, cuando Charles Dickens colocó un Megalosaurus en el primer capítulo de su novela Bleak House en 1852. Los dinosaurios presentados en libros, películas, programas de televisión, obras de arte y otros medios se han utilizado tanto para educación como para entretenimiento. . Las representaciones van desde las realistas, como en los documentales televisivos de la década de 1990 y la primera década del siglo XXI, o las fantásticas, como en las películas de monstruos de los años cincuenta y sesenta.
La serpiente o serpiente ha jugado un poderoso papel simbólico en diferentes culturas. En la historia egipcia, la cobra del Nilo adornaba la corona del faraón. Fue adorado como uno de los dioses y también fue usado con propósitos siniestros: asesinato de un adversario y suicidio ritual (Cleopatra). En la mitología griega, las serpientes se asocian con antagonistas mortales, como un símbolo ctónico, más o menos traducido como terrestre. La Hidra Lernaean de nueve cabezas derrotada por Hércules y las tres hermanas Gorgona son hijos de Gaia, la tierra. Medusa fue una de las tres hermanas Gorgonas que Perseo derrotó. Medusa es descrita como una horrible mortal, con serpientes en vez de pelo y el poder de convertir a los hombres en piedras con su mirada. Después de matarla, Perseo le dio la cabeza a Atenea, quien la fijó en su escudo llamado Aegis. Los Titanes están representados en el arte con sus piernas reemplazadas por cuerpos de serpientes por la misma razón: son hijos de Gaia, por lo que están atados a la tierra. En el hinduismo, las serpientes son adoradas como dioses, y muchas mujeres vierten leche en fosas de serpientes. La cobra se ve en el cuello de Shiva, mientras que Vishnu se representa a menudo como durmiendo en una serpiente de siete cabezas o dentro de las espirales de una serpiente. Hay templos en India únicamente para cobras a veces llamados Nagraj (Rey de las Serpientes), y se cree que las serpientes son símbolos de fertilidad. En el festival hindú anual de Nag Panchami, las serpientes son veneradas y oradas. En términos religiosos, la serpiente y el jaguar son posiblemente los animales más importantes en la antigua Mesoamérica. "En estados de éxtasis, los señores bailan una danza de serpientes, grandes serpientes descendentes adornan y sostienen edificios desde Chichén Itzá hasta Tenochtitlan, y la palabra náhuatl coatl que significa serpiente o gemelo, forma parte de deidades primarias como Mixcoatl, Quetzalcóatl y Coatlicue". En el cristianismo y el judaísmo, una serpiente aparece en Génesis para tentar a Adán y Eva con el fruto prohibido del Árbol del conocimiento del bien y del mal.
La tortuga tiene una posición destacada como símbolo de constancia y tranquilidad en religión, mitología y folclore de todo el mundo. La longevidad de una tortuga es sugerida por su larga vida y su caparazón, que se pensaba que lo protegía de cualquier enemigo. En los mitos cosmológicos de varias culturas, una Tortuga Mundial lleva el mundo sobre su espalda o apoya los cielos.
Medicina

Las muertes por mordeduras de serpientes son poco comunes en muchas partes del mundo, pero todavía se cuentan en decenas de miles por año en la India. Snakebite se puede tratar con antiveneno hecho del veneno de la serpiente. Para producir antiveneno, una mezcla de los venenos de diferentes especies de serpientes se inyecta en el cuerpo de un caballo en dosis cada vez mayores hasta que el caballo se inmuniza. La sangre luego se extrae; el suero se separa, se purifica y se liofiliza. El efecto citotóxico del veneno de serpiente se está investigando como un tratamiento potencial para el cáncer.
Los gecos también se han utilizado como medicina, especialmente en China.
Otro
Los cocodrilos están protegidos en muchas partes del mundo y se cultivan comercialmente. Sus pieles se curten y se utilizan para hacer artículos de cuero como zapatos y bolsos de mano; la carne de cocodrilo también se considera un manjar. Las especies más comúnmente cultivadas son los cocodrilos de agua salada y del Nilo. La agricultura ha resultado en un aumento en la población de cocodrilos de agua salada en Australia, ya que los huevos generalmente se cosechan en la naturaleza, por lo que los propietarios tienen un incentivo para conservar su hábitat. El cuero de cocodrilo está hecho en carteras, carteras, bolsos, bolsos, cinturones, sombreros y zapatos. El aceite de cocodrilo ha sido utilizado para diversos fines.
En el mundo occidental, algunas serpientes (especialmente especies dóciles como la pitón de bola y la serpiente de maíz) se mantienen como mascotas.
Obtenido de: https://en.wikipedia.org/wiki/Reptile