Seleccion natural
Definición

La selección natural es la supervivencia diferencial y la reproducción de los individuos debido a las diferencias en el fenotipo. Es un mecanismo clave de la evolución, el cambio en los rasgos hereditarios característicos de una población a lo largo de generaciones. Charles Darwin popularizó el término "selección natural", contrastándolo con la selección artificial, que es intencional, mientras que la selección natural no lo es.
La variación existe dentro de todas las poblaciones de organismos. Esto ocurre en parte porque surgen mutaciones aleatorias en el genoma de un organismo individual, y la descendencia puede heredar tales mutaciones. A lo largo de la vida de las personas, sus genomas interactúan con sus entornos para causar variaciones en los rasgos. El entorno de un genoma incluye la biología molecular en la célula, otras células, otros individuos, poblaciones, especies, así como el entorno abiótico. Debido a que los individuos con ciertas variantes del rasgo tienden a sobrevivir y a reproducirse más que los individuos con otras variantes menos exitosas, la población evoluciona. Otros factores que afectan el éxito reproductivo incluyen la selección sexual (ahora a menudo incluida en la selección natural) y la selección de la fecundidad.
La selección natural actúa sobre el fenotipo, las características del organismo que realmente interactúan con el ambiente, pero la base genética (hereditaria) de cualquier fenotipo que le da a ese fenotipo una ventaja reproductiva puede volverse más común en una población. Con el tiempo, este proceso puede resultar en poblaciones que se especializan en nichos ecológicos particulares (microevolución) y eventualmente puede resultar en especiación (aparición de nuevas especies, macroevolución). En otras palabras, la selección natural es un proceso clave en la evolución de una población.
La selección natural es una piedra angular de la biología moderna. El concepto, publicado por Darwin y Alfred Russel Wallace en una presentación conjunta de artículos en 1858, fue elaborado en el influyente libro de 1859 sobre el origen de las especies por medio de la selección natural de Darwin, o la preservación de las razas favorecidas en la lucha por la vida. Él describió la selección natural como análoga a la selección artificial, un proceso mediante el cual los animales y plantas con características consideradas deseables por los mejoradores humanos son sistemáticamente favorecidos para la reproducción. El concepto de selección natural desarrollado originalmente en ausencia de una teoría válida de la herencia; En el momento de la escritura de Darwin, la ciencia todavía tenía que desarrollar teorías modernas de la genética. La unión de la evolución darwiniana tradicional con descubrimientos posteriores en genética clásica formó la síntesis moderna de mediados del siglo XX. La incorporación de la genética molecular ha llevado a la biología evolutiva del desarrollo, lo que explica la evolución a nivel molecular. Mientras que los genotipos pueden cambiar lentamente por deriva genética aleatoria, la selección natural sigue siendo la principal explicación para la evolución adaptativa.
Desarrollo historico
Teorías Predarwinianas

Varios filósofos de la era clásica, incluyendo a Empédocles y su sucesor intelectual, el poeta romano Lucrecio, expresaron la idea de que la naturaleza produce una gran variedad de criaturas, al azar, y que solo persisten las criaturas que logran mantenerse y reproducirse con éxito. La idea de Empédocles de que los organismos surgieron enteramente por el funcionamiento incidental de causas como el calor y el frío fue criticada por Aristóteles en el Libro II de la Física . Él postuló la teleología natural en su lugar, y creyó que la forma se logró con un propósito, citando la regularidad de la herencia en las especies como prueba. Sin embargo, aceptó en su biología que los nuevos tipos de animales, monstruosidades (τερας), pueden ocurrir en casos muy raros ( Generación de Animales, Libro IV). Como se cita en la edición de El origen de las especies de Darwin de 1872 , Aristóteles consideró si las diferentes formas (por ejemplo, de los dientes) podrían haber aparecido accidentalmente, pero solo sobrevivieron las formas útiles:
Pero Aristóteles rechazó esta posibilidad en el párrafo siguiente, dejando en claro que está hablando sobre el desarrollo de animales como embriones con la frase "ya sea invariable o normalmente", no el origen de las especies:
La lucha por la existencia fue descrita más tarde por el escritor islámico Al-Jahiz en el siglo IX.
Los argumentos clásicos fueron reintroducidos en el siglo XVIII por Pierre Louis Maupertuis y otros, incluido el abuelo de Darwin, Erasmus Darwin.
Hasta principios del siglo XIX, la opinión predominante en las sociedades occidentales era que las diferencias entre los individuos de una especie eran desviaciones poco interesantes de sus ideales platónicos (o typus) de tipos creados. Sin embargo, la teoría del uniformitarismo en geología promovió la idea de que las fuerzas simples y débiles podrían actuar continuamente durante largos períodos de tiempo para producir cambios radicales en el paisaje de la Tierra. El éxito de esta teoría aumentó la conciencia de la vasta escala del tiempo geológico e hizo plausible la idea de que pequeños cambios, virtualmente imperceptibles en sucesivas generaciones, podrían producir consecuencias en la escala de las diferencias entre las especies.
El zoólogo de principios del siglo XIX Jean-Baptiste Lamarck sugirió la herencia de las características adquiridas como un mecanismo para el cambio evolutivo; los rasgos adaptativos adquiridos por un organismo durante su vida podrían ser heredados por la progenie de ese organismo, eventualmente causando transmutación de especies. Esta teoría, lamarckismo, fue una influencia en el antagonismo del biólogo soviético Trofim Lysenko a la teoría genética dominante hasta mediados del siglo XX.
Entre 1835 y 1837, el zoólogo Edward Blyth trabajó en el área de la variación, la selección artificial y cómo un proceso similar ocurre en la naturaleza. Darwin reconoció las ideas de Blyth en el primer capítulo sobre la variación de Sobre el origen de las especies .
La teoría de Darwin
En 1859, Charles Darwin expuso su teoría de la evolución por selección natural como una explicación para la adaptación y la especiación. Definió la selección natural como el "principio por el cual se preserva cada pequeña variación [de un rasgo], si es útil". El concepto era simple pero poderoso: los individuos mejor adaptados a sus entornos tienen más probabilidades de sobrevivir y reproducirse. Siempre que haya alguna variación entre ellos y que la variación sea hereditaria, habrá una selección inevitable de individuos con las variaciones más ventajosas. Si las variaciones son hereditarias, el éxito reproductivo diferencial conduce a una evolución progresiva de poblaciones particulares de una especie, y las poblaciones que evolucionan para ser lo suficientemente diferentes eventualmente se convierten en especies diferentes.

Las ideas de Darwin se inspiraron en las observaciones que había hecho en el segundo viaje del HMS Beagle (1831-1836) y en el trabajo de un economista político, Thomas Robert Malthus, quien, en Un ensayo sobre el principio de la población (1798), notaron que la población (si no se controla) aumenta exponencialmente, mientras que el suministro de alimentos crece solo aritméticamente; por lo tanto, las limitaciones inevitables de los recursos tendrían implicaciones demográficas, lo que llevaría a una "lucha por la existencia". Cuando Darwin leyó a Malthus en 1838, ya estaba preparado por su trabajo como naturalista para apreciar la "lucha por la existencia" en la naturaleza. Se le ocurrió que a medida que la población aumentaba sus recursos, "las variaciones favorables tendían a conservarse, y las desfavorables a destruirse. El resultado de esto sería la formación de nuevas especies". Darwin escribió:
Una vez que tuvo su teoría, Darwin fue meticuloso a la hora de recopilar y refinar pruebas antes de hacer pública su idea. Estaba en el proceso de escribir su "gran libro" para presentar su investigación cuando el naturalista Alfred Russel Wallace concibió independientemente el principio y lo describió en un ensayo que envió a Darwin para transmitirlo a Charles Lyell. Lyell y Joseph Dalton Hooker decidieron presentar su ensayo junto con escritos inéditos que Darwin había enviado a otros naturalistas, y Sobre la Tendencia de las Especies para formar Variedades; y sobre la Perpetuación de Variedades y Especies por Medios Naturales de Selección fue leída a la Sociedad Linneana de Londres, anunciando el descubrimiento conjunto del principio en julio de 1858. Darwin publicó un relato detallado de su evidencia y conclusiones en Sobre el origen de las especies en 1859. En la 3ª edición de 1861, Darwin reconoció que otros -como William Charles Wells en 1813 y Patrick Matthew en 1831- habían propuesto ideas similares, pero no las habían desarrollado ni presentado en notables publicaciones científicas.

Darwin pensó en la selección natural por analogía con la forma en que los agricultores seleccionan los cultivos o el ganado para la cría, lo que denominó "selección artificial"; en sus primeros manuscritos se refería a una "Naturaleza" que haría la selección. En ese momento, otros mecanismos de evolución como la evolución por deriva genética aún no se habían formulado explícitamente, y Darwin creía que la selección era probablemente solo una parte de la historia: "Estoy convencido de que la selección natural ha sido el principal pero no exclusivo medio de modificación. " En una carta a Charles Lyell en septiembre de 1860, Darwin lamentó el uso del término "Selección Natural", prefiriendo el término "Preservación Natural".
Para Darwin y sus contemporáneos, la selección natural era en esencia sinónimo de evolución por selección natural. Después de la publicación de Sobre el origen de las especies , las personas educadas generalmente aceptaron que la evolución había ocurrido de alguna forma. Sin embargo, la selección natural seguía siendo controvertida como mecanismo, en parte porque se percibía que era demasiado débil para explicar el rango de características observadas de los organismos vivos, y en parte porque incluso los partidarios de la evolución se resistían a su naturaleza "no guiada" y no progresiva, una respuesta que ha sido caracterizada como el impedimento más importante para la aceptación de la idea. Sin embargo, algunos pensadores adoptaron con entusiasmo la selección natural; después de leer a Darwin, Herbert Spencer presentó la frase supervivencia del más apto, que se convirtió en un resumen popular de la teoría. La quinta edición de Sobre el origen de las especies, publicada en 1869, incluyó la frase de Spencer como alternativa a la selección natural, con el mérito de: "Pero la expresión utilizada a menudo por el Sr. Herbert Spencer de la Supervivencia del más apto es más precisa, y algunas veces igualmente conveniente ". Aunque la frase todavía es utilizada a menudo por no biólogos, los biólogos modernos la evitan porque es tautológica si se dice que "más apto" significa "funcionalmente superior" y se aplica a individuos en lugar de considerarse como una cantidad promediada sobre las poblaciones.
La síntesis moderna
La selección natural se basa fundamentalmente en la idea de la herencia, pero se desarrolló antes que los conceptos básicos de la genética. Aunque el Moravianmonk Gregor Mendel, el padre de la genética moderna, era contemporáneo de Darwin, su obra yacía en la oscuridad, solo fue redescubierta en 1900. Con la integración de la evolución de principios del siglo XX con las leyes de herencia de Mendel, la llamada síntesis moderna , los científicos en general llegaron a aceptar la selección natural. La síntesis creció a partir de avances en diferentes campos. Ronald Fisher desarrolló el lenguaje matemático requerido y escribió The Genetical Theory of Natural Selection (1930). JBS Haldane introdujo el concepto del "costo" de la selección natural. Sewall Wright dilucidó la naturaleza de la selección y la adaptación. En su libro La genética y el origen de las especies (1937), Theodosius Dobzhansky estableció la idea de que la mutación, una vez vista como un rival para la selección, en realidad suministraba la materia prima para la selección natural al crear diversidad genética.
Una segunda síntesis

Ernst Mayr reconoció la importancia clave del aislamiento reproductivo para la especiación en su Sistemática y el origen de las especies (1942). WD Hamilton concibió la selección de parentesco en 1964. Esta síntesis consolidó la selección natural como el fundamento de la teoría de la evolución, donde permanece hoy. A fines del siglo XX, se produjo una segunda síntesis por los avances en genética molecular, creando el campo de la biología evolutiva del desarrollo ("evo-devo"), que busca explicar la evolución de la forma en términos de los programas de regulación genética que controlar el desarrollo del embrión a nivel molecular. Se entiende aquí que la selección natural actúa sobre el desarrollo embrionario para cambiar la morfología del cuerpo adulto.
Terminología
El término selección natural generalmente se define para operar en rasgos heredables, porque estos participan directamente en la evolución. Sin embargo, la selección natural es "ciega" en el sentido de que los cambios en el fenotipo pueden dar una ventaja reproductiva independientemente de si el rasgo es hereditario o no. Siguiendo el uso primario de Darwin, el término se usa para referirse tanto a la consecuencia evolutiva de la selección ciega como a sus mecanismos. A veces es útil distinguir explícitamente entre los mecanismos de selección y sus efectos; cuando esta distinción es importante, los científicos definen la "selección natural (fenotípica)" específicamente como "aquellos mecanismos que contribuyen a la selección de individuos que se reproducen", independientemente de si la base de la selección es hereditaria. seleccionado para , mientras que aquellos que reducen el éxito son seleccionados en contra .
Mecanismo
Variación heredable, reproducción diferencial

La variación natural ocurre entre los individuos de cualquier población de organismos. Algunas diferencias pueden mejorar las posibilidades de que un individuo sobreviva y se reproduzca de tal manera que su tasa reproductiva de por vida aumenta, lo que significa que deja más descendencia. Si los rasgos que le dan a estos individuos una ventaja reproductiva también son heredables, es decir, pasados de padres a hijos, habrá una reproducción diferencial, es decir, una proporción ligeramente mayor de conejos rápidos o algas eficientes en la próxima generación. Incluso si la ventaja reproductiva es muy leve, a lo largo de muchas generaciones cualquier rasgo hereditario ventajoso se vuelve dominante en la población. De esta forma, el entorno natural de un organismo "selecciona para" rasgos que confieren una ventaja reproductiva, causando un cambio evolutivo, como describió Darwin. Esto da la apariencia de un propósito, pero en la selección natural no hay una elección intencional. La selección artificial es intencional donde la selección natural no lo es, aunque los biólogos a menudo usan un lenguaje teleológico para describirla.
La polilla moteada existe en colores claros y oscuros en Gran Bretaña, pero durante la revolución industrial, muchos de los árboles en los que descansaban las polillas se ennegrecieron por el hollín, dando a las polillas de color oscuro una ventaja para esconderse de los depredadores. Esto dio a las polillas de color oscuro una mejor oportunidad de sobrevivir para producir crías de color oscuro, y en solo cincuenta años desde la captura de la primera polilla oscura, casi todas las polillas del Manchester industrial estaban a oscuras. El equilibrio fue revertido por el efecto de la Ley de Aire Limpio de 1956, y las polillas oscuras se volvieron raras nuevamente, demostrando la influencia de la selección natural en la evolución de la polilla moteada.
Aptitud
El concepto de aptitud es fundamental para la selección natural. En términos generales, las personas que están más "en forma" tienen un mejor potencial de supervivencia, como en la conocida frase "supervivencia del más apto", pero el significado preciso del término es mucho más sutil. La teoría evolutiva moderna define la aptitud no por cuánto tiempo vive un organismo, sino por el éxito que tiene en la reproducción. Si un organismo vive la mitad de tiempo que otros de su especie, pero tiene el doble de descendencia que sobrevive hasta la edad adulta, sus genes se vuelven más comunes en la población adulta de la próxima generación. Aunque la selección natural actúa sobre individuos, los efectos de la casualidad significan que la aptitud física solo puede definirse realmente "en promedio" para los individuos dentro de una población.
Competencia
En biología, la competencia es una interacción entre organismos en la que la aptitud de uno se ve disminuida por la presencia de otro. Esto puede deberse a que ambos dependen de un suministro limitado de un recurso como alimentos, agua o territorio. La competencia puede ser dentro o entre especies, y puede ser directa o indirecta. Las especies menos adecuadas para competir deberían, en teoría, adaptarse o extinguirse, ya que la competencia desempeña un papel importante en la selección natural, pero según la teoría del "espacio para vagar" puede ser menos importante que la expansión entre clados más grandes.
La competencia se basa en la teoría de la selección de r / K , que se basa en Robert MacArthur y en el trabajo de EO Wilson sobre biogeografía de islas. En esta teoría, las presiones selectivas impulsan la evolución en una de dos direcciones estereotipadas: r - o K -selection. Estos términos, r y K , se pueden ilustrar en un modelo logístico de dinámica de poblaciones:
donde r es la tasa de crecimiento de la población ( N ), y K es la capacidad de carga de su entorno ambiental local. Típicamente, las especies seleccionadas r explotan nichos vacíos y producen muchos descendientes, cada uno con una probabilidad relativamente baja de sobrevivir hasta la edad adulta. En contraste, las especies K seleccionadas son competidores fuertes en nichos atestados, e invierten más en mucha menos descendencia, cada una con una probabilidad relativamente alta de sobrevivir hasta la edad adulta.
Tipos de selección

La selección natural puede actuar sobre cualquier rasgo fenotípico hereditario, y la presión selectiva puede ser producida por cualquier aspecto del medio ambiente, incluida la selección sexual y la competencia con miembros de la misma especie u otras especies. Sin embargo, esto no implica que la selección natural sea siempre direccional y resulte en una evolución adaptativa; la selección natural a menudo resulta en el mantenimiento del status quo al eliminar variantes menos aptas.
La selección puede clasificarse de diferentes maneras, por ejemplo, por su efecto en un rasgo, en la diversidad genética, por la etapa del ciclo de vida en que actúa, por la unidad de selección o por el recurso que se compite.
La selección tiene diferentes efectos en los rasgos. La selección de estabilización actúa para mantener un rasgo en un óptimo estable, y en el caso más simple todas las desviaciones de este óptimo son selectivamente desventajosas. La selección direccional favorece los valores extremos de un rasgo. La selección disruptiva poco común también actúa durante períodos de transición cuando el modo actual es subóptimo, pero altera el rasgo en más de una dirección. En particular, si el rasgo es cuantitativo y univariado, se favorecen los niveles de rasgo más altos y más bajos. La selección disruptiva puede ser un precursor de la especiación.
Alternativamente, la selección se puede dividir según su efecto sobre la diversidad genética. La selección purificadora o negativa actúa para eliminar la variación genética de la población (y se opone a la mutación de novo , que introduce nuevas variaciones. Por el contrario, la selección de equilibrio actúa para mantener la variación genética en una población, incluso en ausencia de una mutación de novo , selección negativa dependiente de la frecuencia. Un mecanismo para esto es la ventaja de heterocigotos, donde individuos con dos alelos diferentes tienen una ventaja selectiva sobre individuos con un solo alelo. El polimorfismo en el locus del grupo sanguíneo ABO humano se ha explicado de esta manera.
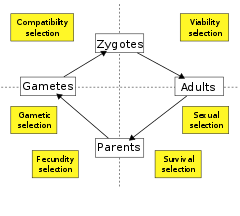
Otra opción es clasificar la selección por la etapa del ciclo de vida en la que actúa. Algunos biólogos reconocen solo dos tipos: selección de viabilidad (o supervivencia), que actúa para aumentar la probabilidad de supervivencia de un organismo y selección de fecundidad (o fertilidad o reproducción), que actúa para aumentar la tasa de reproducción, dada la supervivencia. Otros dividen el ciclo de vida en otros componentes de selección. Así, la selección de viabilidad y supervivencia puede definirse por separado y respectivamente para mejorar la probabilidad de supervivencia antes y después de alcanzar la edad reproductiva, mientras que la selección de fecundidad puede dividirse en subcomponentes adicionales que incluyen selección sexual, selección de gametos, supervivencia del gameto, y selección de compatibilidad, actuando sobre la formación de cigotos.
La selección también se puede clasificar por nivel o unidad de selección. La selección individual actúa sobre el individuo, en el sentido de que las adaptaciones son "para" el beneficio del individuo, y el resultado de la selección entre individuos. La selección de genes actúa directamente al nivel del gen. En la selección de parentesco y el conflicto intragenómico, la selección del nivel del gen proporciona una explicación más adecuada del proceso subyacente. La selección grupal, si ocurre, actúa sobre grupos de organismos, bajo la suposición de que los grupos se replican y mutan de forma análoga a los genes y las personas. Existe un debate continuo sobre el grado en que la selección grupal ocurre en la naturaleza.
Finalmente, la selección se puede clasificar según el recurso para el que se compite. La selección sexual resulta de la competencia por parejas. La selección sexual normalmente se realiza a través de la selección de fecundidad, a veces a expensas de la viabilidad. La selección ecológica es la selección natural por cualquier medio que no sea la selección sexual, como la selección de parentesco, la competencia y el infanticidio. Siguiendo a Darwin, la selección natural a veces se define como selección ecológica, en cuyo caso la selección sexual se considera un mecanismo separado.
Selección sexual

La selección sexual como la articuló primero Darwin (usando el ejemplo de la cola del pavo real) se refiere específicamente a la competencia por parejas, que pueden ser intrasexuales , entre individuos del mismo sexo, es decir, competencia hombre-hombre o intersexual , donde un género elige parejas , lo más a menudo posible con hombres que exhiben y mujeres que eligen. Sin embargo, en algunas especies, la elección de pareja es principalmente por los machos, como en algunos peces de la familia Syngnathidae.
Los rasgos fenotípicos pueden mostrarse en un sexo y desearse en el otro sexo, causando un ciclo de retroalimentación positiva llamado fugitivo de un pescador, por ejemplo, el plumaje extravagante de algunos pájaros macho como el pavo real. Una teoría alternativa propuesta por el mismo Ronald Fisher en 1930 es la hipótesis del hijo sexy, que las madres quieren hijos promiscuos para darles un gran número de nietos y así elegir padres promiscuos para sus hijos. La agresión entre miembros del mismo sexo a veces se asocia con características muy distintivas, como las astas de los ciervos, que se usan en combate con otros ciervos. De manera más general, la selección intrasexual a menudo se asocia con el dimorfismo sexual, incluidas las diferencias en el tamaño corporal entre los machos y las hembras de una especie.
Selección natural en acción
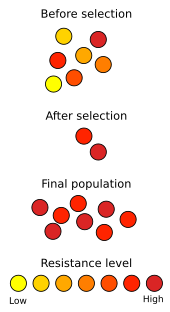
La selección natural se ve en acción en el desarrollo de resistencia a antibióticos en microorganismos. Desde el descubrimiento de la penicilina en 1928, los antibióticos se han usado para combatir las enfermedades bacterianas. El uso indebido generalizado de antibióticos ha seleccionado la resistencia microbiana a los antibióticos en el uso clínico, hasta el punto de que el Staphylococcus aureus resistente a la meticilina (MRSA) ha sido descrito como una "superbacteria" debido a la amenaza que representa para la salud y su relativa invulnerabilidad a las drogas existentes. Las estrategias de respuesta generalmente incluyen el uso de antibióticos diferentes y más fuertes; sin embargo, recientemente han aparecido nuevas cepas de SARM resistentes incluso a estos medicamentos. Esta es una carrera armamentista evolutiva, en la que las bacterias desarrollan cepas menos susceptibles a los antibióticos, mientras que los investigadores médicos intentan desarrollar nuevos antibióticos que puedan matarlos. Una situación similar ocurre con la resistencia a pesticidas en plantas e insectos. Las carreras de brazos no son necesariamente inducidas por el hombre; un ejemplo bien documentado implica la propagación de un gen en la mariposa Hypolimnas bolina que suprime la actividad de matanza masculina por Wolbachiaparásitos bacterianos en la isla de Samoa, donde se sabe que la propagación del gen se produjo en un período de solo cinco años
Evolución mediante selección natural

Un requisito previo para que la selección natural resulte en la evolución adaptativa, los rasgos novedosos y la especiación es la presencia de una variación genética hereditaria que da lugar a diferencias de aptitud física. La variación genética es el resultado de mutaciones, recombinaciones genéticas y alteraciones en el cariotipo (el número, forma, tamaño y disposición interna de los cromosomas). Cualquiera de estos cambios puede tener un efecto que sea altamente ventajoso o muy desventajoso, pero los efectos grandes son raros. En el pasado, la mayoría de los cambios en el material genético se consideraban neutrales o casi neutros porque ocurrían en el ADN no codificante o daban como resultado una sustitución sinónima. Sin embargo, muchas mutaciones en el ADN no codificante tienen efectos perjudiciales. Aunque tanto las tasas de mutación como los efectos promedio de aptitud de las mutaciones dependen del organismo,
Algunas mutaciones ocurren en "kit de herramientas" o genes reguladores. Los cambios en estos a menudo tienen grandes efectos sobre el fenotipo del individuo porque regulan la función de muchos otros genes. La mayoría, pero no todas, las mutaciones en los genes reguladores dan como resultado embriones no viables. Algunas mutaciones reguladoras no letales ocurren en los genes HOX en humanos, lo que puede dar como resultado una polidactilia de Ribor cervical, un aumento en el número de dedos de manos y pies. Cuando tales mutaciones resultan en una mayor aptitud, la selección natural favorece estos fenotipos y el rasgo novedoso se propaga en la población. Los rasgos establecidos no son inmutables; los rasgos que tienen una buena forma física en un contexto ambiental pueden ser mucho menos adecuados si cambian las condiciones ambientales. En ausencia de selección natural para preservar dicho rasgo, se vuelve más variable y se deteriora con el tiempo, posiblemente resultando en una manifestación vestigial del rasgo, también llamado equipaje evolutivo. En muchas circunstancias, la estructura aparentemente vestigial puede retener una funcionalidad limitada, o puede ser cooptada para otros rasgos ventajosos en un fenómeno conocido como preadaptación. Se cree que un famoso ejemplo de estructura vestigial, el ojo de la rata topo ciega, conserva su función en la percepción del fotoperíodo.
Especiación
La especiación requiere un grado de aislamiento reproductivo, es decir, una reducción en el flujo de genes. Sin embargo, es intrínseco al concepto de una especie contra la cual se seleccionan los híbridos, oponiéndose a la evolución del aislamiento reproductivo, un problema que fue reconocido por Darwin. El problema no ocurre en la especiación alopátrica con poblaciones geográficamente separadas, que pueden divergir con diferentes conjuntos de mutaciones. EB Poulton se dio cuenta en 1903 de que el aislamiento reproductivo podría evolucionar por divergencia, si cada linaje adquiriera un alelo diferente e incompatible del mismo gen. La selección contra el heterocigoto crearía un aislamiento reproductivo directo, lo que llevaría al modelo de Bateson-Dobzhansky-Muller, más elaborado por H. Allen Orr y Sergey Gavrilets. Con refuerzo, sin embargo,
Base genética
Genotipo y fenotipo
La selección natural actúa sobre el fenotipo o características físicas de un organismo. El fenotipo está determinado por la composición genética (genotipo) de un organismo y el entorno en el que vive el organismo. Cuando diferentes organismos en una población poseen diferentes versiones de un gen para un determinado rasgo, cada una de estas versiones se conoce como un alelo. Es esta variación genética la que subyace a las diferencias en el fenotipo. Un ejemplo son los antígenos del tipo de sangre ABO en humanos, donde tres alelos gobiernan el fenotipo.
Algunos rasgos están gobernados por un solo gen, pero la mayoría de los rasgos están influenciados por las interacciones de muchos genes. Una variación en uno de los muchos genes que contribuyen a un rasgo puede tener solo un pequeño efecto en el fenotipo; en conjunto, estos genes pueden producir un continuo de posibles valores fenotípicos.
Direccionalidad de la selección
Cuando algún componente de un rasgo es heredable, la selección altera las frecuencias de los diferentes alelos o variantes del gen que produce las variantes del rasgo. La selección se puede dividir en tres clases, en función de su efecto sobre las frecuencias de los alelos: selección direccional, estabilizadora y purificadora. La selección direccional ocurre cuando un alelo tiene una mejor forma física que otros, de modo que aumenta en frecuencia, ganando una participación creciente en la población. Este proceso puede continuar hasta que el alelo sea fijo y la población completa comparta el fenotipo más en forma. Mucho más común es la selección estabilizadora, que disminuye la frecuencia de los alelos que tienen un efecto perjudicial sobre el fenotipo, es decir, producen organismos de menor aptitud física. Este proceso puede continuar hasta que el alelo se elimine de la población.
Algunas formas de selección de equilibrio no resultan en la fijación, pero mantienen un alelo en frecuencias intermedias en una población. Esto puede ocurrir en especies diploides (con pares de cromosomas) cuando los individuos heterocigotos (con solo una copia del alelo) tienen una aptitud más alta que los individuos homocigotos (con dos copias). Esto se conoce como ventaja heterocigota o sobredominio, de los cuales el ejemplo más conocido es la resistencia a la malaria en humanos heterocigotos para la anemia de células falciformes. El mantenimiento de la variación alélica también puede ocurrir a través de una selección disruptiva o diversificadora, lo que favorece los genotipos que se apartan del promedio en cualquier dirección (es decir, lo opuesto al exceso de dominancia) y puede dar como resultado una distribución bimodal de los valores de los rasgos. Finalmente, la selección de equilibrio puede ocurrir a través de la selección dependiente de la frecuencia, donde la aptitud de un fenotipo particular depende de la distribución de otros fenotipos en la población. Los principios de la teoría de juegos se han aplicado para comprender las distribuciones de la aptitud en estas situaciones, particularmente en el estudio de la selección de parentesco y la evolución del altruismo recíproco.
Selección, variación genética y deriva
Una porción de toda variación genética es funcionalmente neutral, no produce ningún efecto fenotípico o diferencia significativa en la aptitud. La teoría neutra de Motoo Kimura sobre la evolución molecular por deriva genética propone que esta variación represente una gran fracción de la diversidad genética observada. Los eventos neutrales pueden reducir radicalmente la variación genética a través de los cuellos de botella de la población. que entre otras cosas puede causar el efecto fundador en inicialmente pequeñas poblaciones nuevas. Cuando la variación genética no da lugar a diferencias en la aptitud, la selección no puede afectar directamente la frecuencia de dicha variación. Como resultado, la variación genética en esos sitios es más alta que en los sitios donde la variación influye en la aptitud física. Sin embargo, después de un período sin nuevas mutaciones, la variación genética en estos sitios se elimina debido a la deriva genética. La selección natural reduce la variación genética al eliminar individuos inadaptados y, en consecuencia, las mutaciones que causaron la inadaptación. Al mismo tiempo, ocurren nuevas mutaciones, que dan como resultado un equilibrio de selección de mutación. El resultado exacto de los dos procesos depende tanto de la velocidad a la que ocurren las nuevas mutaciones como de la fuerza de la selección natural, que es una función de cuán desfavorable resulta la mutación.
La vinculación genética ocurre cuando los loci de dos alelos están muy cerca de un cromosoma. Durante la formación de los gametos, la recombinación reorganiza los alelos. La posibilidad de que tal reorganización ocurra entre dos alelos está inversamente relacionada con la distancia entre ellos. Los barridos selectivos se producen cuando un alelo se vuelve más común en una población como resultado de una selección positiva. A medida que aumenta la prevalencia de un alelo, los alelos estrechamente vinculados también pueden volverse más comunes mediante el "autoestop genético", ya sean neutrales o incluso levemente perjudiciales. Un barrido selectivo fuerte da como resultado una región del genoma donde el haplotipo seleccionado positivamente (el alelo y sus vecinos) son, en esencia, los únicos que existen en la población. Los barridos selectivos se pueden detectar midiendo el desequilibrio de ligamiento, o si un haplotipo dado está sobrerrepresentado en la población. Como un barrido selectivo también da como resultado la selección de alelos vecinos, la presencia de un bloque de fuerte desequilibrio de ligamiento podría indicar un barrido selectivo "reciente" cerca del centro del bloque.
La selección de fondo es lo opuesto a un barrido selectivo. Si un sitio específico experimenta una selección de purificación fuerte y persistente, la variación vinculada tiende a eliminarse junto con ella, produciendo una región en el genoma de baja variabilidad general. Debido a que la selección de fondo es el resultado de nuevas mutaciones perjudiciales, que pueden ocurrir al azar en cualquier haplotipo, no produce bloques claros de desequilibrio de ligamiento, aunque con baja recombinación todavía puede conducir a un desequilibrio de ligamiento ligeramente negativo en general.
Impacto
Las ideas de Darwin, junto con las de Adam Smith y Karl Marx, tuvieron una profunda influencia en el pensamiento del siglo XIX, incluyendo su afirmación radical de que "formas elaboradamente construidas, tan diferentes entre sí y dependientes entre sí de una manera tan compleja" evolucionaron desde las formas más simples de vida mediante unos simples principios. Esto inspiró a algunos de los partidarios más ardientes de Darwin y provocó la oposición más fuerte. La selección natural tenía el poder, según Stephen Jay Gould, de "destronar algunas de las comodidades más profundas y tradicionales del pensamiento occidental", como la creencia de que los seres humanos tienen un lugar especial en el mundo.
En palabras del filósofo Daniel Dennett, "la peligrosa idea de Darwin" de la evolución por selección natural es un "ácido universal", que no puede mantenerse restringido a ningún recipiente o recipiente, ya que pronto se filtra, abriéndose paso en un camino cada vez más amplio. alrededores. Así, en las últimas décadas, el concepto de selección natural se ha extendido desde la biología evolutiva a otras disciplinas, incluidas la computación evolutiva, el darwinismo cuántico, la economía evolutiva, la epistemología evolutiva, la psicología evolutiva y la selección cosmológica natural. Esta aplicabilidad ilimitada ha sido llamada darwinismo universal.
Origen de la vida
Cómo la vida se originó a partir de la materia inorgánica sigue siendo un problema no resuelto en la biología. Una hipótesis prominente es que la vida apareció por primera vez en forma de polímeros de ARN autoreplicantes cortos. Desde este punto de vista, la vida pudo haber existido cuando las cadenas de ARN experimentaron por primera vez las condiciones básicas, tal como las concibió Charles Darwin, para que la selección natural funcione. Estas condiciones son: heredabilidad, variación de tipo y competencia por recursos limitados. La aptitud de un replicador de ARN temprano probablemente habría sido función de las capacidades de adaptación que eran intrínsecas (es decir, determinadas por la secuencia de nucleótidos) y la disponibilidad de recursos. Las tres capacidades principales de adaptación podrían haber sido lógicamente: (1) la capacidad de replicarse con fidelidad moderada (lo que da lugar a heredabilidad y variación de tipo), (2) la capacidad de evitar la descomposición, y (3) la capacidad de adquirir y procesar recursos. Estas capacidades se habrían determinado inicialmente por las configuraciones plegadas (incluidas aquellas configuraciones con actividad de ribozima) de los replicadores de ARN que, a su vez, se habrían codificado en sus secuencias de nucleótidos individuales.
Célula y biología molecular
En 1881, el embriólogo Wilhelm Roux publicó Der Kampf der Theile im Organismus ( La lucha de las partes en el organismo ) en el que sugirió que el desarrollo de un organismo resulta de una competencia darwiniana entre las partes del embrión, que se produce en todos los niveles, de las moléculas a los órganos. En los últimos años, Jean-Jacques Kupiec ha propuesto una versión moderna de esta teoría. De acuerdo con este Darwinismo celular, la variación aleatoria a nivel molecular genera diversidad en los tipos de células mientras que las interacciones celulares imponen un orden característico en el embrión en desarrollo.
Teoría social y psicológica
Las implicaciones sociales de la teoría de la evolución por selección natural también se convirtieron en fuente de continua controversia. Friedrich Engels, un filósofo político alemán y coautor de la ideología del comunismo, escribió en 1872 que "Darwin no sabía qué amarga sátira escribió a la humanidad, y especialmente a sus compatriotas, cuando demostró que la libre competencia, la lucha para la existencia, que los economistas celebran como el mayor logro histórico, es el estado normal del reino animal. "Herbert Spencer y el defensor eugenésico Francis Galton interpretan la selección natural como necesariamente progresiva, lo que lleva a supuestos avances en inteligencia y civilización, se convirtió en una justificación para el colonialismo, eugenesia y darwinismo social. Por ejemplo, en 1940, Konrad Lorenz, en escritos que posteriormente repudió, usó la teoría como una justificación para las políticas del estado nazi. Escribió: "... la selección de la dureza, el heroísmo y la utilidad social ... debe ser realizada por alguna institución humana, si la humanidad, en defecto de factores selectivos, no debe ser arruinado por la degeneración inducida por la domesticación. La idea racial como la base de nuestro estado ya ha logrado mucho en este sentido ".
Más recientemente, el trabajo entre antropólogos y psicólogos ha llevado al desarrollo de la sociobiología y más tarde de la psicología evolutiva, un campo que intenta explicar las características de la psicología humana en términos de adaptación al entorno ancestral. El ejemplo más prominente de la psicología evolutiva, notablemente avanzado en los primeros trabajos de Noam Chomsky y más tarde por Steven Pinker, es la hipótesis de que el cerebro humano se ha adaptado para adquirir las reglas gramaticales del lenguaje natural. Se ha formulado la hipótesis de que otros aspectos del comportamiento humano y las estructuras sociales, desde normas culturales específicas, como la evitación del incesto hasta patrones más amplios, como los roles de género, tienen orígenes similares a las adaptaciones al entorno primitivo en el que evolucionaron los humanos modernos. Por analogía con la acción de la selección natural sobre los genes, el concepto de memes ...
Teoría de información y sistemas
En 1922, Alfred J. Lotka propuso que la selección natural podría entenderse como un principio físico que podría describirse en términos del uso de la energía por un sistema, un concepto desarrollado más tarde por Howard T. Odum como el principio de máxima potencia en termodinámica, mediante el cual los sistemas evolutivos con ventaja selectiva maximizan la tasa de transformación de energía útil.
Los principios de la selección natural han inspirado una variedad de técnicas computacionales, como la vida artificial "suave", que simula procesos selectivos y puede ser altamente eficiente en entidades 'adaptoras' a un ambiente definido por una función específica de aptitud. Por ejemplo, una clase de algoritmos de optimización heurística conocidos como algoritmos genéticos, iniciados por John Henry Holland en la década de 1970 y ampliados por David E. Goldberg, identifican soluciones óptimas por reproducción simulada y mutación de una población de soluciones definidas por una distribución de probabilidad inicial . Tales algoritmos son particularmente útiles cuando se aplican a problemas cuyo paisaje energético es muy desigual o tiene muchos mínimos locales.
En ficción
La evolución darwiniana por selección natural es omnipresente en la literatura, ya sea optimista en términos de cómo la humanidad puede evolucionar hacia la perfección, o pesimista en términos de las nefastas consecuencias de la interacción de la naturaleza humana y la lucha por la supervivencia. Entre las principales respuestas se encuentra Erewhon pesimista 1872 de Samuel Butler ("en ninguna parte", escrito al revés). En 1893 HG Wells imaginó "El Hombre del Año Millón", transformado por selección natural en un ser con una enorme cabeza y ojos, y cuerpo encogido.
Obtenido de: https://en.wikipedia.org/wiki/Natural_selection