Semilla
Definición

Una semilla es una planta embrionaria encerrada en una cubierta exterior protectora. La formación de la semilla es parte del proceso de reproducción en las plantas de semillas, los espermatofitos, incluidas las plantas de gimnospermas y angiospermas.
Las semillas son el producto del óvulo maduro, después de la fertilización por polen y un poco de crecimiento dentro de la planta madre. El embrión se desarrolla a partir del cigoto y la cubierta de la semilla de las epidermis del óvulo.
Las semillas han sido un desarrollo importante en la reproducción y el éxito de las plantas de gimnospermas y angiospermas, en relación con las plantas más primitivas, como helechos, musgos y hepáticas, que no tienen semillas y utilizan medios dependientes del agua para propagarse. Las plantas de semillas ahora dominan los nichos biológicos en la tierra, desde los bosques hasta las praderas tanto en climas cálidos como fríos.
El término "semilla" también tiene un significado general que es anterior a lo anterior: cualquier cosa que se pueda sembrar, por ejemplo, papas "semilla", "semillas" de "semillas" de maíz o girasol. En el caso de las "semillas" de girasol y maíz, lo que se siembra es la semilla encerrada en una cáscara o cáscara, mientras que la papa es un tubérculo.
Muchas estructuras comúnmente conocidas como "semillas" son en realidad frutos secos. Las plantas que producen bayas se llaman baccate. Las semillas de girasol a veces se venden comercialmente mientras están encerradas dentro de la pared dura de la fruta, que debe dividirse para alcanzar la semilla. Los diferentes grupos de plantas tienen otras modificaciones, las llamadas frutas de hueso (como el melocotón) tienen una capa de fruta endurecida (el endocarpio) fusionada a la semilla real y que la rodea. Las nueces son los frutos de una sola semilla y cáscara dura de algunas plantas con una semilla indehiscente, como una bellota o avellana.
Producción de semillas
Las semillas se producen en varios grupos relacionados de plantas, y su modo de producción distingue las angiospermas ("semillas cerradas") de las gimnospermas ("semillas desnudas"). Las semillas de angiospermas se producen en una estructura dura o carnosa llamada fruta que encierra las semillas para protegerlas a fin de asegurar un crecimiento saludable. Algunas frutas tienen capas de material duro y carnoso. En las gimnospermas, no se desarrolla una estructura especial para encerrar las semillas, que comienzan su desarrollo "desnudas" en las brácteas de los conos. Sin embargo, las semillas quedan cubiertas por las escamas del cono a medida que se desarrollan en algunas especies de coníferas.
La producción de semillas en las poblaciones de plantas naturales varía ampliamente de un año a otro en respuesta a variables climáticas, insectos y enfermedades, y ciclos internos dentro de las propias plantas. En un período de 20 años, por ejemplo, los bosques compuestos por pino de esquisto y pino de hoja corta produjeron de 0 a casi 5 millones de semillas de pino sanas por hectárea. Durante este período, se evaluaron seis cosechas abundantes, cinco pobres y nueve buenas, cuando se evaluó la producción de plántulas adecuadas para la reproducción del bosque natural.
Desarrollo
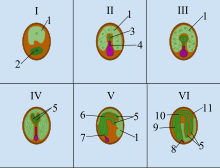
Las semillas de angiospermas (plantas con flores) constan de tres componentes genéticamente distintos: (1) el embrión formado a partir del cigoto, (2) el endospermo, que normalmente es triploide, (3) el revestimiento de la semilla del tejido derivado del tejido materno del óvulo . En las angiospermas, el proceso de desarrollo de la semilla comienza con la doble fertilización, que implica la fusión de dos gametos masculinos con el óvulo y la célula central para formar el endosper primario y el zigoto. Justo después de la fecundación, el cigoto es en su mayoría inactivo, pero el endospermo primario se divide rápidamente para formar el tejido del endosperma. Este tejido se convierte en el alimento que la planta joven consumirá hasta que las raíces se hayan desarrollado después de la germinación.
Óvulo

Después de la fertilización, los óvulos se desarrollan en las semillas. El óvulo consiste en una serie de componentes:
- El funículo ( funículo, funículo ) o tallo de la semilla que une el óvulo a la placenta y, por lo tanto, el ovario o la pared de la fruta, en el pericarpio .
- El nucelo , el remanente del megasporangio y la región principal del óvulo donde se desarrolla el megagametofito.
- El micropilo , un poro pequeño o abertura en el ápice de la epidermis del óvulo donde el tubo de polen generalmente entra durante el proceso de fertilización.
- La chalaza , la base del óvulo opuesto al micropilo, donde el integumento y el nucelo se unen.
La forma de los óvulos a medida que se desarrollan a menudo afecta la forma final de las semillas. Las plantas generalmente producen óvulos de cuatro formas: la forma más común se llama anátropo , con una forma curva. Los óvulos ortotópicos son rectos con todas las partes del óvulo alineadas en una fila larga produciendo una semilla no curvada. Los óvulos Campylotropous tienen un megagametofito curvo que a menudo le da a la semilla una forma apretada de "C". La última forma de óvulo se llama amphitropous , donde el óvulo está parcialmente invertido y vuelto hacia atrás 90 grados en su tallo (el funículo o funículo ).
En la mayoría de las plantas con flores, la primera división del cigoto está orientada transversalmente con respecto al eje largo, y esto establece la polaridad del embrión. El polo superior o chalazal se convierte en el área principal de crecimiento del embrión, mientras que el polo inferior o micropilar produce el suspensor en forma de tallo que se adhiere al micropilo. El suspensor absorbe y fabrica nutrientes del endosperma que se usan durante el crecimiento del embrión.
Embrión

Los principales componentes del embrión son:
- Los cotiledones , las hojas de la semilla, unidas al eje embrionario. Puede haber uno (Monocotiledóneas) o dos (Dicotiledóneas). Los cotiledones también son la fuente de nutrientes en las dicotiledóneas no endospérmicas, en cuyo caso reemplazan al endosperma y son gruesas y correosas. En las semillas endospérmicas, los cotiledones son delgados y de papel. Los dicotiledóneos tienen el punto de unión uno frente al otro en el eje.
- El epicotilo , el eje embrionario por encima del punto de unión del cotiledón (es).
- La plúmula , la punta del epicotilo, y tiene una apariencia plumosa debido a la presencia de primordios foliares jóvenes en el ápice, y se convertirá en el brote en la germinación.
- El hipocotilo , el eje embrionario debajo del punto de unión del cotiledón (es), conectando el epicotilo y la radícula, que es la zona de transición del tallo-raíz.
- La radícula , la punta basal del hipocótilo, crece en la raíz primaria.
Las plantas monocotiledóneas tienen dos estructuras adicionales en forma de vainas. La plúmula se cubre con un coleoptilo que forma la primera hoja, mientras que la radícula se cubre con una coleorhiza que se conecta con la raíz primaria y las raíces adventicias se forman desde los lados. Aquí el hipocótilo es un eje rudimentario entre la radícula y la plúmula. Las semillas de maíz se construyen con estas estructuras; pericarpio, escutelo (único cotiledón grande) que absorbe nutrientes del endospermo, plúmula, radícula, coleoptilo y coleorhiza; estas dos últimas estructuras son similares a vainas y encierran la plúmula y la radícula, actuando como una cubierta protectora.
Cubierta de semilla
El óvulo en maduración experimenta cambios marcados en las epidermis, generalmente una reducción y desorganización pero ocasionalmente un engrosamiento. La cubierta de la semilla se forma a partir de los dos tegumentos o las capas externas de las células del óvulo, que se derivan de los tejidos de la planta madre, el integumento interior forma el tegmen y el exterior forma la testa. (Las capas de semillas de algunas plantas mononocotiledóneas, como los pastos, no son estructuras distintas, sino que se fusionan con la pared de la fruta para formar un pericarpio). Los testados de monocotiledóneas y dicotiledóneas a menudo están marcados con patrones y marcas texturizadas, o alas o mechones de cabello. Cuando la cubierta de la semilla se forma a partir de una sola capa, también se denomina testa, aunque no todas las testae son homólogas de una especie a la siguiente. El funículo abscisa (se separa en el punto fijo - zona de abscisión), la cicatriz forma una depresión oval, el hilio . Los óvulos anatrópodos tienen una porción del funículo que es adnato (fusionado a la cubierta de la semilla), y que forma una cresta longitudinal, o rafe , justo arriba del hilio. En óvulos bitegámicos (p. Ej., Gossypium descrito aquí), tanto los integumentos internos como los externos contribuyen a la formación del recubrimiento de la semilla. Con la maduración continua, las células se agrandan en el integumento externo. Mientras que la epidermis interna puede permanecer como una sola capa, también puede dividirse para producir dos o tres capas y acumular almidón, y se conoce como la capa incolora. Por el contrario, la epidermis externa se vuelve tanífera. El integumento interno puede consistir en ocho a quince capas. (Kozlowski 1972)
A medida que las células se agrandan y el almidón se deposita en las capas externas de la zona pigmentada debajo de la epidermis externa, esta zona comienza a lignificarse, mientras que las células de la epidermis externa se agrandan radialmente y sus paredes se engrosan, con núcleo y citoplasma comprimidos en el exterior capa. estas células que son más amplias en su superficie interna se llaman células de empalizada . En la epidermis interna, las células también se agrandan radialmente con un engrosamiento similar a la placa de las paredes. El integumento interno maduro tiene una capa de empalizada, una zona pigmentada con 15-20 capas, mientras que la capa más interna se conoce como la capa marginal. (Kozlowski 1972)
Gimnospermas
En las gimnospermas, que no forman ovarios, los óvulos y, por lo tanto, las semillas están expuestos. Esta es la base de su nomenclatura: plantas desnudas con semillas. Dos espermatozoides transferidos desde el polen no desarrollan la semilla por fertilización doble, pero un núcleo de espermatozoides se une con el núcleo del huevo y el otro esperma no se usa.
Algunas veces cada esperma fertiliza un óvulo y un cigoto es luego abortado o absorbido durante el desarrollo temprano. La semilla está compuesta por el embrión (el resultado de la fertilización) y el tejido de la planta madre, que también forma un cono alrededor de la semilla en plantas coníferas como el pino y el abeto.
Forma y apariencia
Se usa una gran cantidad de términos para describir las formas de las semillas, muchas de las cuales se explican por sí mismas, como en forma de frijol (reniforme), parecido a un riñón, con extremos lobulados a cada lado del hilio, cuadrada u oblonga , angular con todas lados más o menos iguales o más largos que anchos, Triangulares : tres lados, más ancho debajo del medio, elípticos u ovados u ovalados , redondeados en ambos extremos, o con forma de huevo (ovados u obovados, más anchos en un extremo), redondeados pero simétricos el medio o más ancho debajo del medio o más ancho sobre el medio.
Otros términos menos obvios incluyen discoide (que se asemeja a un disco o placa, que tiene tanto grosor como caras paralelas y con un margen redondeado), elipsoide , globoso (esférico) o subgloboso (inflado, pero menos que esférico), lenticular , oblongo , ovoide , reniforme y sectoroide . Estriar las semillas tienen rayas paralelas, líneas longitudinales o crestas. Los colores más comunes son marrón y negro, otros colores son poco frecuentes. La superficie varía de altamente pulida a considerablemente rugosa. La superficie puede tener una variedad de apéndices (ver Capa de semilla). Una capa de semilla con la consistencia del corcho se conoce como suberosa. Otros términos incluyen crustáceos (duros, delgados o frágiles).
Estructura

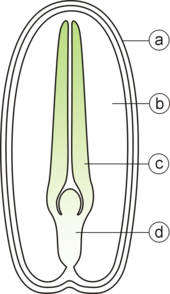
Una semilla típica incluye dos partes básicas:
- un embrión;
- una capa de semilla.
Además, el endosperma forma un suministro de nutrientes para el embrión en la mayoría de las monocotiledóneas y las dicotiledóneas endospérmicas.
Tipos de semillas
Se ha considerado que las semillas ocurren en muchos tipos estructuralmente diferentes (Martin 1946). Estos se basan en una serie de criterios, de los cuales el dominante es la relación de tamaño de embrión a semilla. Esto refleja el grado en que los cotiledones en desarrollo absorben los nutrientes del endospermo y, por lo tanto, lo destruyen.
Seis tipos ocurren entre las monocotiledóneas, diez en las dicotiledóneas y dos en las gimnospermas (lineales y espatuladas). Esta clasificación se basa en tres características: la morfología del embrión, la cantidad de endosperma y la posición del embrión en relación con el endospermo.

Embrión
En las semillas endospérmicas, hay dos regiones distintas dentro de la cubierta de la semilla, un endosperma superior y un embrión más pequeño y más pequeño. El embrión es el óvulo fertilizado, una planta inmadura de la cual una nueva planta crecerá en las condiciones adecuadas. El embrión tiene un cotiledón o una hoja semillera en monocotiledones, dos cotiledones en casi todas las dicotiledóneas y dos o más en gimnospermas. En el fruto de los granos (cariópsides), la única monocotiledónea tiene forma de escudo y, por lo tanto, se llama escutelo . El escutelo se presiona estrechamente contra el endosperma del cual absorbe los alimentos y lo pasa a las partes en crecimiento. Las descripciones de embriones incluyen pequeñas, rectas, dobladas, curvas y rizadas.
Almacenamiento de nutrientes
Dentro de la semilla, generalmente hay una reserva de nutrientes para la plántula que crecerá a partir del embrión. La forma de la nutrición almacenada varía según el tipo de planta. En las angiospermas, el alimento almacenado comienza como un tejido llamado endospermo, que se deriva de la planta madre y el polen a través de la doble fertilización. Por lo general, es triploide y es rico en aceite o almidón y proteínas. En las gimnospermas, como las coníferas, el tejido de almacenamiento de alimentos (también llamado endospermo) es parte del gametofito femenino, un tejido haploide. El endospermo está rodeado por la capa de aleurona (endosperma periférico), lleno de granos de aleurona proteínica.
Originalmente, por analogía con el óvulo animal, la capa externa del nucelo (perisperma) se denominaba albúmina, y la capa interna del endospermo como vitelo. Aunque engañoso, el término comenzó a aplicarse a toda la materia nutritiva. Esta terminología persiste al referirse a las semillas endospérmicas como "albuminosas". La naturaleza de este material se utiliza tanto en la descripción como en la clasificación de las semillas, además de la relación entre el tamaño del embrión y el endosperma. El endospermo puede considerarse farináceo (o harinoso) en el que las células se llenan con almidón, como por ejemplo granos de cereal, o no (no farináceas). El endosperma también puede denominarse "carnoso" o "cartilaginoso" con células blandas más gruesas como el coco, pero también puede ser aceitoso como en Ricinus (aceite de ricino), Croton y Poppy. El endospermo se denomina "córneo" cuando las paredes de las células son más gruesas, como la fecha y el café, o "rumiado" si está moteado, como en nuez moscada, palmas y Annonaceae.
En la mayoría de las monocotiledóneas (como las gramíneas y las palmas) y algunas dicotiledóneas ( endospérmicas o albuminosas ) (como el ricino), el embrión está incrustado en el endospermo (y nucelo), que la plántula usará en la germinación. En las dicotiledóneas no endospérmicas, el embrión absorbe el endospermo a medida que éste crece dentro de la semilla en desarrollo, y los cotiledones del embrión se llenan con alimentos almacenados. En la madurez, las semillas de estas especies no tienen endospermo y también se las conoce como exalbuminous semillas Las semillas exalbuminous incluyen las legumbres (tales como habas y guisantes), árboles tales como el roble y el nogal, vehículos tales como calabaza y rábano, y girasoles. Según Bewley y Black (1978), el almacenamiento de la nuez de Brasil está en el hipocótilo, este lugar de almacenamiento es poco común entre las semillas. Todas las semillas de gimnosperma son albuminosas.
Cubierta de semilla
La cubierta de la semilla se desarrolla a partir del tejido materno, las epidermis, que originalmente rodeaban el óvulo. La cubierta de la semilla madura puede ser una capa delgada como el papel (por ejemplo, maní) o algo más sustancial (por ejemplo, espeso y duro en la pulpa de la miel y el coco), o carnoso como en la sarcotesta de la granada. La cubierta de la semilla ayuda a proteger al embrión de lesiones mecánicas, depredadores y resequedad. Dependiendo de su desarrollo, la cubierta de la semilla es bien bitegumentado o unitegmic. Las semillas bitegmicas forman una testa desde el integumento externo y un tegmen desde el integumento interno, mientras que las semillas unitérmicas tienen una sola membrana. Por lo general, partes de la testa o tegmen forman una capa mecánica protectora dura. La capa mecánica puede evitar la penetración del agua y la germinación. Entre las barreras puede estar la presencia de esclereidos lignificados.
El integumento externo tiene varias capas, generalmente entre cuatro y ocho organizadas en tres capas: (a) epidermis externa, (b) zona pigmentada externa de dos a cinco capas que contienen tanino y almidón, y (c) epidermis interna. El endotegmen se deriva de la epidermis interna del integumento interno, el exotegmen de la superficie externa del integumento interno. La endotesta se deriva de la epidermis interna del integumento externo, y la capa externa de la testa de la superficie externa del tegumento exterior se conoce como la exotesta. Si el exotesta es también la capa mecánica, esto se llama una semilla exotestal, pero si la capa mecánica es el endotegmen, entonces la semilla es endotestal. La exotesta puede consistir en una o más filas de células que son alargadas y palidecidas (por ejemplo, Fabaceae), por lo tanto, "empalizada exotesta".
Además de las tres partes básicas de semillas, algunas semillas tienen un apéndice, un arilo , un crecimiento carnoso del funículo (funículo), (como en el tejo y la nuez moscada) o un apéndice graso, un elaiosoma (como en Corydalis ), o pelos (tricomas). En el último ejemplo, estos pelos son la fuente del algodón de cultivo textil. Otros apéndices de semillas incluyen el rafe (una cresta), alas, carúnculas (un crecimiento suave y esponjoso del epitelio externo en las proximidades del micropilo), espinas o tubérculos.
También puede quedar una cicatriz en la cubierta de la semilla, llamada hilio , donde el funículo ataca la semilla a la pared del ovario. Justo debajo de él hay un poro pequeño, que representa el micropilo del óvulo.
Tamaño y conjunto de semillas

Las semillas son muy diversas en tamaño. Las semillas de orquídeas similares al polvo son las más pequeñas, con alrededor de un millón de semillas por gramo; a menudo son semillas embrionarias con embriones inmaduros y sin reservas energéticas significativas. Las orquídeas y algunos otros grupos de plantas son mycoheterotrophs que dependen de los hongos micorrízicos para la nutrición durante la germinación y el crecimiento temprano de la plántula. Algunas plantas de orquídeas terrestres, de hecho, pasan los primeros años de sus vidas obteniendo energía de los hongos y no producen hojas verdes. Con más de 20 kg, la semilla más grande es el coco de mer. Las plantas que producen semillas más pequeñas pueden generar muchas más semillas por flor, mientras que las plantas con semillas más grandes invierten más recursos en esas semillas y normalmente producen menos semillas. Las semillas pequeñas maduran más rápidamente y se pueden dispersar antes, por lo que las plantas que caen en floración a menudo tienen semillas pequeñas. Muchas plantas anuales producen grandes cantidades de semillas más pequeñas; esto ayuda a asegurar que al menos unos pocos terminarán en un lugar favorable para el crecimiento. Las plantas perennes herbáceas y las plantas leñosas a menudo tienen semillas más grandes; pueden producir semillas durante muchos años, y las semillas más grandes tienen más reservas de energía para la germinación y el crecimiento de plántulas y producen plántulas más grandes y más establecidas después de la germinación.
Funciones
Las semillas cumplen varias funciones para las plantas que las producen. Las principales funciones son la nutrición del embrión, la dispersión a una nueva ubicación y la latencia en condiciones desfavorables. Las semillas son fundamentalmente medios de reproducción, y la mayoría de las semillas son el producto de la reproducción sexual que produce una remezcla de material genético y la variabilidad fenotípica en la que actúa la selección natural.
Alimento embrionario
Las semillas protegen y nutren el embrión o la planta joven. Por lo general, dan a las plántulas un comienzo más rápido que las esporas de una espora, debido a las mayores reservas de alimentos en la semilla y la multicelularidad del embrión incluido.
Dispersión
A diferencia de los animales, las plantas tienen una capacidad limitada para buscar condiciones favorables para la vida y el crecimiento. Como resultado, las plantas han desarrollado muchas formas de dispersar a su descendencia mediante la dispersión de sus semillas (véase también la reproducción vegetativa). Una semilla debe de alguna manera "llegar" a un lugar y estar allí en un momento favorable para la germinación y el crecimiento. Cuando los frutos se abren y liberan sus semillas de manera regular, se llama dehiscente, que a menudo es distintivo para grupos de plantas relacionados; estas frutas incluyen cápsulas, folículos, legumbres, siliconas y silicuas. Cuando las frutas no se abren y liberan sus semillas de manera regular, se llaman indehiscentes, que incluyen los frutos aquenio, cariópside, nueces, samaras y utrículos.
Por el viento (anemochory)


- Algunas semillas (p. Ej., Pino) tienen un ala que ayuda a la dispersión del viento.
- Las semillas de orquídeas como polvo son transportadas eficientemente por el viento.
- Algunas semillas (p. Ej. Algodoncillo, álamo) tienen pelos que ayudan a la dispersión del viento.
Otras semillas están encerradas en estructuras frutales que ayudan a la dispersión del viento de manera similar:
- Los aquenios de diente de león tienen pelos.
- Las samaras de arce tienen dos alas.
Por agua (hidrochory)
- Algunas plantas, como Mucuna y Dioclea , producen semillas vigorosas llamadas alubias o semillas de deriva porque flotan en los ríos hasta los océanos y se lavan en las playas.
Por animales (zoochory)
- Semillas (rebabas) con púas o ganchos (por ejemplo, acaena, bardana, muelle) que se adhieren a la piel o plumas de animales, y luego se dejan más tarde.
- Las semillas con una cubierta carnosa (por ejemplo, manzana, cereza, enebro) son comidas por animales (aves, mamíferos, reptiles, peces) que luego dispersan estas semillas en sus excrementos.
- Las semillas (frutos secos) son recursos alimenticios almacenables atractivos a largo plazo para los animales (por ejemplo, bellotas, avellanas, nueces); las semillas se almacenan a cierta distancia de la planta progenitora, y algunas escapan para ser comidas si el animal las olvida.
Myrmecochory es la dispersión de las semillas por las hormigas. Las hormigas forrajeras dispersan semillas que tienen apéndices llamados elaiosomas (p. Ej., Sanguinaria, trilliums, acacias y muchas especies de Proteaceae). Los elaiosomas son estructuras suaves y carnosas que contienen nutrientes para los animales que los comen. Las hormigas llevan tales semillas a su nido, donde se comen los elaiosomas. El resto de la semilla, que es dura y no comestible para las hormigas, luego germina dentro del nido o en un sitio de remoción donde las hormigas han desechado la semilla. Esta relación de dispersión es un ejemplo de mutualismo, ya que las plantas dependen de las hormigas para dispersar las semillas, mientras que las hormigas dependen de las semillas de las plantas para obtener alimento. Como resultado, una disminución en el número de un socio puede reducir el éxito del otro. En Sudáfrica, la hormiga argentina (Linepithema humile ) ha invadido y desplazado especies nativas de hormigas. A diferencia de las especies de hormigas nativas, las hormigas argentinas no recolectan las semillas de Mimetes cucullatus ni comen los elaiosomas. En las áreas donde estas hormigas han invadido, el número de plántulas de Mimetes ha disminuido.
Inactividad
La latencia de las semillas tiene dos funciones principales: la primera es sincronizar la germinación con las condiciones óptimas para la supervivencia de la plántula resultante; el segundo es difundir la germinación de un lote de semillas a lo largo del tiempo, por lo que una catástrofe (por ejemplo, heladas tardías, sequía, herbivoría) no da como resultado la muerte de todos los descendientes de una planta (cobertura de apuestas). La latencia de las semillas se define como una semilla que no germina en condiciones ambientales óptimas para la germinación, normalmente cuando el ambiente está a una temperatura adecuada con la humedad adecuada del suelo. Esta verdadera latencia o inactividad innata es causada por las condiciones dentro de la semilla que impiden la germinación. Por lo tanto, la latencia es un estado de la semilla, no del medio ambiente. Latencia inducida
La latencia de las semillas no es lo mismo que la persistencia de las semillas en el suelo o en la planta, aunque incluso en las publicaciones científicas, la latencia y la persistencia a menudo se confunden o se usan como sinónimos.
Con frecuencia, la latencia de la semilla se divide en cuatro categorías principales: exógena; endógeno; combinacional; y secundario Un sistema más reciente distingue cinco clases: latencia morfológica, fisiológica, morfofisiológica, física y combinacional.
La latencia exógena es causada por afecciones fuera del embrión, que incluyen:
- La dormancia física o las capas duras de las semillas se producen cuando las semillas son impermeables al agua. En la pausa de latencia, una estructura especializada, la "brecha de agua", se interrumpe en respuesta a señales ambientales, especialmente la temperatura, por lo que el agua puede entrar en la semilla y puede ocurrir la germinación. Las familias de plantas donde ocurre latencia física incluyen Anacardiaceae, Cannaceae, Convulvulaceae, Fabaceae y Malvaceae.
- La latencia química considera especies que carecen de latencia fisiológica, pero donde un producto químico impide la germinación. Este químico puede ser lixiviado de la semilla por el agua de lluvia o nieve derretida o puede ser desactivado de alguna manera. La filtración de inhibidores químicos de la semilla por el agua de lluvia a menudo se cita como una causa importante de liberación de latencia en las semillas de plantas del desierto, pero existen pocas pruebas para apoyar esta afirmación.
La latencia endógena es causada por afecciones dentro del embrión en sí, que incluyen:
- En la latencia morfológica , se previene la germinación debido a las características morfológicas del embrión. En algunas especies, el embrión es solo una masa de células cuando las semillas están dispersas; no está diferenciado Antes de que se produzca la germinación, deben ocurrir tanto la diferenciación como el crecimiento del embrión. En otras especies, el embrión se diferencia, pero no se cultiva completamente (subdesarrolla) en la dispersión, y se requiere el crecimiento del embrión hasta una longitud específica de la especie antes de que pueda ocurrir la germinación. Ejemplos de familias de plantas donde ocurre latencia morfológica son Apiaceae, Cycadaceae, Liliaceae, Magnoliaceae y Ranunculaceae.
- La latencia morfofisiológica incluye semillas con embriones subdesarrollados, y también tienen componentes fisiológicos para la latencia. Estas semillas, por lo tanto, requieren tratamientos de interrupción de latencia, así como un período de tiempo para desarrollar embriones completamente desarrollados. Las familias de plantas donde ocurre latencia morfofisiológica incluyen Apiaceae, Aquifoliaceae, Liliaceae, Magnoliaceae, Papaveraceae y Ranunculaceae. Algunas plantas con latencia morfofisiológica, como Asarum o Trillium especies, tienen múltiples tipos de latencia, uno afecta el crecimiento de la radícula (raíz), mientras que el otro afecta el crecimiento de la plúmula (brote). Los términos "doble dormancia" y "semillas de dos años" se utilizan para las especies cuyas semillas necesitan dos años para completar la germinación o al menos dos inviernos y un verano. La latencia de la radícula (raíz de la plántula) se rompe durante el primer invierno después de la dispersión, mientras que la latencia del brote se rompe durante el segundo invierno.
- Latencia fisiológica significa que el embrión, debido a causas fisiológicas, no puede generar la potencia suficiente para atravesar el recubrimiento de la semilla, el endosperma u otras estructuras de cobertura. La latencia generalmente se rompe en condiciones de frío, humedad, calor húmedo o calor seco. El ácido abscísico suele ser el inhibidor del crecimiento en las semillas, y su producción puede verse afectada por la luz.
- El secado , en algunas plantas, incluyendo varias hierbas y aquellas de regiones estacionalmente áridas, es necesario antes de que germinen. Las semillas se liberan, pero deben tener un contenido de humedad menor antes de que pueda comenzar la germinación. Si las semillas permanecen húmedas después de la dispersión, la germinación se puede retrasar durante muchos meses o incluso años. Muchas plantas herbáceas de las zonas climáticas templadas tienen latencia fisiológica que desaparece con el secado de las semillas. Otras especies germinarán después de la dispersión solo bajo rangos de temperatura muy estrechos, pero a medida que las semillas se secan, pueden germinar en un rango de temperatura más amplio.
- En las semillas con latencia combinacional , la semilla o la capa de fruta es impermeable al agua y el embrión tiene latencia fisiológica. Dependiendo de la especie, la latencia física se puede romper antes o después de que se rompe la latencia fisiológica.
- La latencia secundaria * es causada por condiciones después de que la semilla se ha dispersado y ocurre en algunas semillas cuando la semilla no libre está expuesta a condiciones que no son favorables para la germinación, muy a menudo a altas temperaturas. Los mecanismos de latencia secundaria aún no se comprenden por completo, pero pueden implicar la pérdida de sensibilidad en los receptores en la membrana plasmática.
Los siguientes tipos de latencia de la semilla no involucran la latencia de la semilla, hablando estrictamente, ya que la falta de germinación es prevenida por el ambiente, no por las características de la misma semilla (ver Germinación):
- La fotodormancia o la sensibilidad a la luz afectan la germinación de algunas semillas. Estas semillas fotoblásticas necesitan un período de oscuridad o luz para germinar. En especies con capas finas de semillas, la luz puede penetrar en el embrión dormido. La presencia de luz o la ausencia de luz pueden desencadenar el proceso de germinación, inhibiendo la germinación en algunas semillas enterradas demasiado profundamente o en otras no enterradas en el suelo.
- La termodormancia es la sensibilidad de la semilla al calor o al frío. Algunas semillas, como el berberecho y el amaranto, germinan solo a altas temperaturas (30 ° C o 86 ° F); muchas plantas que tienen semillas que germinan temprano en pleno verano tienen termodormancia, por lo que germinan solo cuando la temperatura del suelo es cálida. Otras semillas necesitan suelos fríos para germinar, mientras que otras, como el apio, se inhiben cuando la temperatura del suelo es demasiado cálida. A menudo, los requisitos de termodormancia desaparecen a medida que la semilla envejece o se seca.
No todas las semillas experimentan un período de latencia. Las semillas de algunos manglares son vivíparas; comienzan a germinar mientras todavía están unidos al padre. La raíz grande y pesada permite que la semilla penetre en el suelo cuando cae. Muchas semillas de plantas de jardín germinarán fácilmente tan pronto como tengan agua y estén lo suficientemente calientes; aunque sus antepasados salvajes pudieron haber dormido, estas plantas cultivadas carecen de él. Después de muchas generaciones de presión selectiva por parte de los fitomejoradores y los jardineros, se seleccionó la latencia.
Para las plantas anuales, las semillas son una forma para que la especie sobreviva a temporadas secas o frías. Las plantas efímeras suelen ser plantas anuales que pueden pasar de una semilla a otra en tan solo seis semanas.
Persistencia y bancos de semillas
Germinación

La germinación de semillas es un proceso mediante el cual un embrión de semillas se desarrolla en una plántula. Implica la reactivación de las vías metabólicas que conducen al crecimiento y la aparición de la radícula o raíz de la semilla y plúmula o brote. La aparición de la plántula sobre la superficie del suelo es la siguiente fase del crecimiento de la planta y se llama establecimiento de plántulas.
Deben existir tres condiciones fundamentales antes de que pueda ocurrir la germinación. (1) El embrión debe estar vivo, llamado viabilidad de la semilla. (2) Se deben superar los requisitos de latencia que impiden la germinación. (3) Deben existir las condiciones ambientales adecuadas para la germinación.
La viabilidad de las semillas es la capacidad del embrión para germinar y se ve afectada por una serie de condiciones diferentes. Algunas plantas no producen semillas que tengan embriones completos funcionales, o la semilla puede no tener ningún embrión en absoluto, a menudo llamadas semillas vacías. Los depredadores y patógenos pueden dañar o matar a la semilla mientras todavía está en la fruta o después de que se dispersa. Las condiciones ambientales como las inundaciones o el calor pueden matar la semilla antes o durante la germinación. La edad de la semilla afecta su salud y capacidad de germinación: dado que la semilla tiene un embrión vivo, con el tiempo las células mueren y no pueden ser reemplazadas. Algunas semillas pueden vivir durante un tiempo prolongado antes de la germinación, mientras que otras solo pueden sobrevivir durante un breve período después de la dispersión antes de morir.
El vigor de la semilla es una medida de la calidad de la semilla e implica la viabilidad de la semilla, el porcentaje de germinación, la tasa de germinación y la fuerza de las plántulas producidas.
El porcentaje de germinación es simplemente la proporción de semillas que germinan de todas las semillas sujetas a las condiciones adecuadas para el crecimiento. La tasa de germinación es el tiempo que tardan las semillas en germinar. Los porcentajes y tasas de germinación se ven afectados por la viabilidad de la semilla, la latencia y los efectos ambientales que impactan en la semilla y las plántulas. En agricultura y horticultura, las semillas de calidad tienen una alta viabilidad, medida por el porcentaje de germinación más la tasa de germinación. Esto se da como un porcentaje de germinación durante un cierto período de tiempo, 90% de germinación en 20 días, por ejemplo. 'Dormancy' está cubierto arriba; muchas plantas producen semillas con distintos grados de latencia, y diferentes semillas de la misma fruta pueden tener diferentes grados de latencia. Es posible tener semillas sin letargo si se dispersan de inmediato y no se secan (si las semillas se secan entran en latencia fisiológica).
Las condiciones ambientales que afectan la germinación de la semilla incluyen; agua, oxígeno, temperatura y luz.
Se producen tres fases distintas de germinación de la semilla: imbibición del agua; fase de latencia; y la aparición de la radícula.
Para que la cubierta se separe, el embrión debe empaparse (absorber el agua), lo que hace que se hinche, dividiendo la cubierta de la semilla. Sin embargo, la naturaleza de la cubierta de la semilla determina qué tan rápido puede penetrar el agua y posteriormente iniciar la germinación. La tasa de imbibición depende de la permeabilidad de la cubierta de la semilla, la cantidad de agua en el ambiente y el área de contacto de la semilla con la fuente de agua. Para algunas semillas, beber demasiada agua demasiado rápido puede matar la semilla. Para algunas semillas, una vez que se embebe el agua, el proceso de germinación no puede detenerse, y el secado se vuelve fatal. Otras semillas pueden empaparse y perder agua varias veces sin causar efectos nocivos, pero el secado puede causar latencia secundaria.
Reparación de daños en el ADN
Durante la latencia de la semilla, a menudo asociada con ambientes impredecibles y estresantes, el daño del ADN se acumula a medida que las semillas envejecen. En semillas de centeno, la reducción de la integridad del ADN debido al daño se asocia con la pérdida de la viabilidad de la semilla durante el almacenamiento. Tras la germinación, las semillas de Vicia faba se someten a reparación de ADN. Una ADN ligasa vegetal que interviene en la reparación de roturas de cadena simple o doble durante la germinación de la semilla es un determinante importante de la longevidad de la semilla. Además, en las semillas de Arabidopsis, las actividades de las enzimas de reparación del ADN Poly ADP ribosa polimerasas (PARP) son probablemente necesarias para una germinación exitosa. Por lo tanto, los daños en el ADN que se acumulan durante la latencia parecen ser un problema para la supervivencia de la semilla, y la reparación enzimática de los daños del ADN durante la germinación parece ser importante para la viabilidad de la semilla.
Inducir la germinación
Los jardineros y horticultores utilizan una serie de estrategias diferentes para romper la latencia de las semillas.
Escarificación permite que el agua y los gases penetren en la semilla; incluye métodos para romper físicamente las capas de semillas duras o ablandarlas con productos químicos, como remojar en agua caliente o hacer agujeros en la semilla con un alfiler o frotar sobre papel de lija o agrietar con una prensa o un martillo. Algunas veces, los frutos se cosechan mientras las semillas aún están inmaduras y el recubrimiento de la semilla no se desarrolla completamente y se siembra enseguida antes de que la cubierta de la semilla se vuelva impermeable. En condiciones naturales, los roedores mastican las semillas cuando roen la semilla, las semillas rozan las rocas (las semillas son movidas por el viento o las corrientes de agua) al congelarse y descongelarse las aguas superficiales o al pasar por el tracto digestivo de un animal. En este último caso, la cubierta protectora protege la semilla de la digestión, mientras que a menudo debilita el recubrimiento de la semilla de modo que el embrión está listo para brotar cuando se deposita, junto con un poco de materia fecal que actúa como fertilizante, lejos de la planta madre. Los microorganismos a menudo son efectivos para romper las capas duras de las semillas y algunas veces son utilizados por las personas como tratamiento; las semillas se almacenan en un medio arenoso cálido y húmedo durante varios meses bajo condiciones no estériles.
La estratificación , también llamada enfriamiento en húmedo, descompone la latencia fisiológica e implica la adición de humedad a las semillas para que absorban agua, y luego son sometidas a un período de enfriamiento húmedo para madurar el embrión. La siembra a fines del verano y el otoño y la posibilidad de pasar el invierno en condiciones frías es una forma efectiva de estratificar las semillas; algunas semillas responden más favorablemente a períodos de temperaturas oscilantes que son parte del medio ambiente natural.
La lixiviación o remojo en agua elimina los inhibidores químicos en algunas semillas que impiden la germinación. La lluvia y la nieve derretida naturalmente cumplen esta tarea. Para las semillas sembradas en jardines, lo mejor es utilizar agua corriente: si se empapa en un recipiente, es suficiente con remojar de 12 a 24 horas. Remojar más tiempo, especialmente en agua estancada, puede provocar la falta de oxígeno y la muerte de la semilla. Las semillas con capas de semillas duras se pueden remojar en agua caliente para romper las capas de células impermeables que impiden la entrada de agua.
Otros métodos utilizados para ayudar en la germinación de semillas que tienen latencia incluyen preenjuague, presecado, alternancia diaria de temperatura, exposición a la luz, nitrato de potasio, el uso de reguladores del crecimiento de las plantas, como giberelinas, citoquininas, etileno, tiourea, hipoclorito de sodio y otros. Algunas semillas germinan mejor después de un incendio. Para algunas semillas, el fuego raja las capas de las semillas duras, mientras que en otros, la latencia química se rompe como reacción a la presencia de humo. El humo líquido a menudo es utilizado por los jardineros para ayudar en la germinación de estas especies.
Semillas estériles
Las semillas pueden ser estériles por algunas razones: pueden haber sido irradiadas, no haber sido hinchadas, haber sobrevivido a las células o haber sido criadas para tal fin.
Evolución y origen de las semillas
El origen de las plantas de semillas es un problema que aún permanece sin resolver. Sin embargo, cada vez más datos tienden a ubicar este origen en el Devónico medio. La descripción en 2004 de la proto-semilla Runcaria heinzelinii en el Givetian de Bélgica es una indicación de ese antiguo origen de las plantas de semillas. Al igual que con los helechos modernos, la mayoría de las plantas terrestres antes de este tiempo se reprodujeron mediante el envío de esporas al aire, que aterrizarían y se convertirían en plantas completamente nuevas.
Las primeras semillas "verdaderas" se describen desde el Devónico superior, que es probablemente el teatro de su verdadera primera radiación evolutiva. Con esta radiación vino una evolución del tamaño de la semilla, la forma, la dispersión y, finalmente, la radiación de gimnospermas y angiospermas y monocotiledóneas y dicotiledóneas. Las plantas de semillas se convirtieron progresivamente en uno de los principales elementos de casi todos los ecosistemas.
Importancia economica

Semillas comestibles
Muchas semillas son comestibles y la mayoría de las calorías humanas provienen de semillas, especialmente de cereales, legumbres y nueces. Las semillas también proporcionan la mayoría de los aceites de cocina, muchas bebidas y especias y algunos aditivos alimentarios importantes. En diferentes semillas, el embrión de la semilla o el endospermo domina y proporciona la mayoría de los nutrientes. Las proteínas de almacenamiento del embrión y el endospermo difieren en su contenido de aminoácidos y sus propiedades físicas. Por ejemplo, el gluten de trigo, importante para proporcionar la propiedad elástica a la masa de pan, es estrictamente una proteína de endosperma.
Las semillas se utilizan para propagar muchos cultivos, como cereales, legumbres, árboles forestales, céspedes y pastos. Particularmente en los países en desarrollo, una limitación importante que enfrenta es la insuficiencia de los canales de comercialización para llevar la semilla a los agricultores pobres. Por lo tanto, el uso de semilla retenida por los agricultores sigue siendo bastante común.
Las semillas también son comidas por los animales, y se alimentan al ganado. Muchas semillas se usan como alpiste.
Veneno y seguridad alimentaria
Mientras que algunas semillas son comestibles, otras son dañinas, venenosas o mortales. Las plantas y las semillas a menudo contienen compuestos químicos para desalentar a los herbívoros y depredadores de semillas. En algunos casos, estos compuestos simplemente saben mal (como en la mostaza), pero otros compuestos son tóxicos o se descomponen en compuestos tóxicos dentro del sistema digestivo. Los niños, al ser más pequeños que los adultos, son más susceptibles al envenenamiento por las plantas y las semillas.
Un veneno mortal, la ricina, proviene de semillas de ricino. Las dosis letales reportadas oscilan entre dos y ocho semillas, aunque solo se han reportado algunas muertes cuando los animales han ingerido las semillas de ricino.
Además, las semillas que contienen amígdala, manzana, albaricoque, almendra amarga, durazno, ciruela, cereza, membrillo y otras, cuando se consumen en cantidades suficientes, pueden causar envenenamiento por cianuro. Otras semillas que contienen venenos incluyen annona, algodón, chirimoya, datura, durian crudo, cadena de oro, castaño de indias, espuela de caballero, locoweed, lichi, nectarina, rambután, guisante de rosario, soplo agrio, manzana de azúcar, glicina y tejo. Las semillas del árbol de estricnina también son venenosas y contienen la estricnina venenosa.
Las semillas de muchas leguminosas, incluido el frijol común ( Phaseolus vulgaris ), contienen proteínas llamadas lectinas que pueden causar angustia gástrica si los frijoles se comen sin cocinar. El frijol común y muchos otros, incluida la soja, también contienen inhibidores de tripsina que interfieren con la acción de la enzima digestiva tripsina. Los procesos de cocción normales degradan las lectinas y los inhibidores de la tripsina a formas inocuas.
Por favor, consulte la categoría de toxinas vegetales para más artículos relevantes.
Otros usos
La fibra de algodón crece unida a las semillas de plantas de algodón. Otras fibras de semillas son de kapok y milkweed.
Muchos aceites no alimentarios importantes se extraen de las semillas. El aceite de linaza se usa en pinturas. El aceite de jojoba y crambe es similar al aceite de ballena.
Las semillas son la fuente de algunos medicamentos como el aceite de ricino, el aceite de árbol de té y el medicamento contra el cáncer Quack Laetrile.
Muchas semillas se han usado como cuentas en collares y rosarios, incluyendo lágrimas de Job, Chinaberry, guisante de rosario y ricino. Sin embargo, los últimos tres también son venenosos.
Otros usos de semillas incluyen:
- Semillas utilizadas una vez como pesos para saldos.
- Semillas usadas como juguetes por niños, como para el juego Conkers.
- Resina de semillas de Clusia rosea utilizadas para calafatear barcos.
- Nematicida de semillas de algodoncillo.
- Harina de semilla de algodón utilizada como alimento para animales y fertilizante.
Registros de semillas

- La semilla viable más antigua con carbono-14 que ha crecido hasta convertirse en una planta fue una semilla de palmera datilera de Judea de unos 2.000 años de antigüedad, recuperada de las excavaciones en el palacio de Herodes el Grande en Masada, Israel. Fue germinado en 2005. (Una regeneración reportada de Silene stenophylla(Campion de hoja estrecha) de material preservado durante 31.800 años en el permafrost siberiano se logró utilizando tejido de fruta, no de semilla).
- La semilla más grande es producida por el coco de mer, o "doble palma de coco", Lodoicea maldivica . La fruta entera puede pesar hasta 23 kilogramos (50 libras) y generalmente contiene una sola semilla.
- Las semillas más pequeñas son producidas por orquídeas epifitas. Tienen solo 85 micrómetros de largo y pesan 0.81 microgramos. No tienen endospermato y contienen embriones subdesarrollados.
- Las primeras semillas de fósiles tienen alrededor de 365 millones de años del Devónico tardío de Virginia Occidental. Las semillas son óvulos inmaduros preservados de la planta Elkinsia polymorpha .
En religión
El Libro del Génesis en el Antiguo Testamento comienza con una explicación de cómo comenzaron todas las formas de las plantas:
El Corán habla de la germinación de la semilla así: